Membranlar Kimyasal bile im ve organizasyon
advertisement

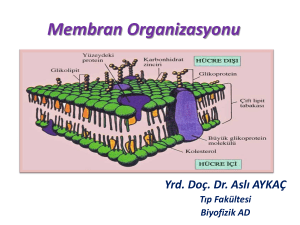
MEMBANLAR: KİMYASAL BİLEŞİM VE ORGANİZASYON Prof.Dr.Hafize Uzun Hücre Membranlarının Özellikleri Biyolojik membranlar hücre içini dış ortamdan ve hücre içi organelleri sitoplazmadan ayıran ileri derecede visköz, fakat esnek yapılardır. Plazma membranı hücrelerin dış sınırlarını belirlemekte ve bu sınırlar içindeki moleküler trafiği düzenlemektedir. İntrasellüler membranlar hücre içi boşluğu, hücresel işlemleri ve elemanları ayırmak üzere kompartmanlara böler ve böylece mitokondri, endoplazmik retikulum, golgi aygıtı, lizozomlar, salgı vesikülleri, nukleus gibi birbirinden yapısal olarak ayırt edilen organelleri oluşturur. Membranlar hücre ve organellere fonksiyonlarına uygun yapı ve şekil kazandırırlar. Hücre Membranlarının Özellikleri Hücresel bütünlük: plazma membranı Hücre içi kompartmanlar: organeller Viskos, plastik, dinamik, asimetrik çift tabaka Lipidler, proteinler, karbonhidratlar Termodinamik olarak stabil Metabolik olarak aktif Nonkovalan oluşum Plazma membranının dış yüzünde peptid hormonlar, nörotransmitterler gibi sinyal molekülleri için spesifik reseptör proteinleri bulunmaktadır.Bu reseptörler hücre dışı sinyalleri hücre içine iletmekte ve hücre içi moleküler değişiklikleri tetiklemektedir. Plazma membranları endositoz ve eksositoz yolu ile dış ortamla madde alışverişi yapmaktadır. Ayrıca birbirine bitişik komşu hücreler arasında aralık kavşaklar(gap junctions) adı verilen spesifik membran yapıları üzerinden madde alışverişi sağlanmaktadır. Membranlar hücre içinde lipid ve belirli proteinlerin sentezi ve mitokondride enerji iletimi gibi hücresel olayları organize etmektedirler. Adezyon molekülleri komşu hücreleri birbirine bağlamaktadır. Plazma membranının dış yüzünde benzer hücrelerin birbirlerini tanımasını ve bir araya gelmesini sağlayan bileşikler bulunmaktadır Membranlar rijid veya impermeabl değil, oldukça mobil ve dinamik yapılardır. Membranların yapısı, organizmalar arası hücreler arası ve organeller arası farklılıklar gösterir Plazma Membranının Fonksiyonları Bariyer Selektif permeabilite Endositoz, eksositoz, “gap junction” Reseptörler ve sinyal iletimi Tanıma ve hücreler arası etkileşim Hücrenin şekil ve hareketi (örn.amiboid hareket) Bazı hücreler yalancı ayaklar(uzantılar) oluşturarak hareketlilik sağlıyabilirler. Bu çıkıntılar ile yabancı maddeleri fagozite ederler. Hareketleri sırasında yer değiştirmeleride mümkündür. Bu hareketlere amiboid hareket denir. Hücre yüzeylerindeki transport hareketleri ve kontraktilite Biyolojik membran yapısı , ‘sıvı akışkan mozaik model ‘ Lipitler ve proteinler membranda, Singer ve Nicolson ( 1972 ) tarafından önerilen, sıvı akışkan mozaik modeli olarak tanımlanan bir düzende yerleşmektedir. Bu modele göre, biyolojik membranların temel yapısını fosfolipid çift tabakası oluşturmakta, Bazı proteinler (integral veya intrinsik proteinler) bu lipid çift tabakayı bir veya birkaç kez katetmekte ve her iki tarafta sulu çevre ile temas etmektedir, diğerleri ise kısmen gömülmüş olarak bulunmaktadır. Bazı proteinler (periferik veya ekstrinsik proteinler) lipid çift tabakanın iç veya dış yüzeyine gevşek bir şekilde bağlı bulunmaktadır Membran lipidleri yapılarında hidrofilik ve hidrofobik grupları içermeleri nedeniyle amfipatik bileşiklerdir. Amfipatik lipidlerin hidrofilik polar baş kısımları su ile ilişkili ve hidrofobik nonpolar kuyruk kısımları (yağ açil zincirleri) ise sudan uzak olan membranın merkezinde bir araya gelecek şekilde yerleşerek lipid çift tabakayı oluştururlar. Biyolojik membranların temel yapısını lipid çift tabakası oluşturmaktadır. Lipid çift tabakanın merkezi iç kısmında bulunan fosfolipid yağ açil zincirleri sıvı, hidrofobik bölge oluşturur. Membran düzlemi üzerinde protein ve lipid moleküllerinin büyük kısmının lateral olarak hareket etmeleri ve bir mozaik yapı oluşturmaları nedeniyle, bu membran yapısına akışkan sıvı-mozaik adı verilmiştir. Membran lipid çift tabakasında yer alan bazı protein ve lipidler kovalent olarak bağlı karbonhidrat zincirleri içerir. Glikoproteinler ve glikolipidlerin bir parçası olan bu karbonhidrat zincirleri daima membranın hücre dışı yüzeyinde, sulu ortama bakacak şekilde asimetrik yerleşim gösterir. Hücrenin dış yüzeyindeki bu karbonhidrat katmanına glikokaliks (hücre mantosu-örtüsü) adı verilir. glikokaliks (hücre mantosu-örtüsü) Hücreyi mekanik hasarlara karşı korur. İstenmeyen protein-protein etkileşimlerinden korur (lektinler). Hücrelerin birbirine yapışmasına hizmet eder. Örneğin kanın pıhtılaşması Membranların kimyasal bileşimleri Membran Protein ( %) Miyelin 20 Eritrosit 49 Karaciğer(p.m) 54 Dış mit. 50 İç mit. 75 Lipid ( %) 75 43 39 46 23 Karbohidrat (%) 5 8 7 4 2 Membran Lipidleri Biyolojik membranların temel yapısını bimoleküler lipid çift tabakası oluşturmaktadır. Lipid çift tabakanın amfipatik özellikteki temel lipid bileşenleri: fosfolipidler (gliserofosfolipidler ve sfingomiyelin), kolesterol glikosfingolipidler ( glikolipidler ) dir. Lipid çift tabakanın oluşumundan hidrofobik etkileşimler sorumludur; Membran lipidlerinin yapısındaki nonpolar hidrokarbon yan zincirlerin bir araya gelmesi suyun uzaklaşmasına yol açar ve hidrokarbon zincirleri van der Waals kuvvetleri ile bir arada tutulur. Lipidlerin polar grupları ile su molekülleri arasında ise elektrostatik etkileşimler ve hidrojen bağları oluşur. Bu şekilde lipid tabakaları non-kovalent etkileşimlerle bir arada tutulur. Fosfolipidler : Fosfogliseridler (gliserofosfolipidler) C16- C18 yağ asidleri, doymuş/doymamış (%50 doymamış), cis Hidrokarbon zinciri kısalırsa hidrofobik karakter ve hacim azalır. Fosfatidik asid Etanolamin, kolin, serin, gliserol ve inositol Fosfatidilkolin(lesitin), fosfatidiletanolamin(sefalin), fosfatidilserin, fosfatidilinozitol ve difosfatidilgliserol (kardiyolipin),plazmalojen Kolin fosfogliseridleri: 16:0 , 18:2 Etanolamin ve serin fosfogliseridler: 20:4 , C22 ÇDYA • Sfingofosfolipidler Sfingomiyelin: seramid, fosforilkolin • Fosfatidilinozitol 4-fosfat ve 4,5-bisfosfat plazma membranlarında bulunur; bunlardan ikincisi “ikinci haberci” olarak tanımlanan ve bazı hormonların etki mekanizmasında yer alan inozitol trifosfat ve diaçilgliserol’ün kaynağıdır. Sinyal iletisi Kardiyolipin (difosfatidilgliserol), plazma membranının yapısında bulunmaz ; az miktarda endoplazmik retikulum membranında buna karşılık yüksek miktarda iç ve daha düşük düzeyde dış mitokondri membranında bulunur. Her bir hücre tipinin membran gliserofosfolipidleri farklı yağ asit bileşimine sahiptir. Ayrıca, gliserofosfolipidlerin yağ asit içeriği dokunun fizyolojik ve patofizyolojik durumuna bağlı olarak değişiklik gösterebilir. Membran akışkanlığını doymuş ve doymamış yağ asitleri güçlü bir şekilde etkilemektedir. Doymuş yağ asitleri düz zincirlidir ve fosfolipid yapısında düz kuyruklar oluşturur. Benzer şekilde doğal olarak bulunmayan ve trans konfigürasyonda çift bağ içeren trans yağ asitleri de düz zincirlidir. Doğal olarak bulunan doymamış yağ asitlerinde çift bağların hemen hepsi cis konfigürasyonundadır ve molekül cis çift bağda bükülmüştür. Bu hidrokarbon zincirlerinde kıvrımlar, kırıklar oluşturur Membran fosfolipidlerinin yapısında doymamış yağ asid miktarı ve doymamışlık derecesi ne kadar fazla ise, fosfolipid kuyruk kısmını oluşturan bu yağ açil zincirleri o kadar çok kıvrım içerir ve membranda o kadar az sıkı paketlenir, böylece membran daha akışkan hal alır. Sfingofosfolipidler Sfingomiyelin: seramid, fosforilkolin Nöron aksonlarının myelin membranlarında çok ! Doymuş ve C24 yağ asidleri • Glikosfingolipidler Serebrozid Serebrozidler, seramid yapısında yer alan sfingozin’in primer alkol grubuna -glikozidik bağ ile bağlanmış bir şeker içermektedirler. Fosfat içermezler, elektriksel olarak nötral glikosfingolipidlerdir Galaktoserebrozid (Galaktozilseramid) beyin ve sinir hücrelerinde çok ! Glukoserebrozidler (Glukozilseramid) Globosit Sülfatid Diglukoz, digalaktoz, N-asetilglukozamin ve N-asetilgalaktozamin başlıca bulunan şekerlerdir. Gangliozid Oligosakkarid zinciri içeren kompleks yapılı, asidik özellikte, Nasetil nöraminik asid içerir. Beyin, sinir dokusunda çobol bulunur. antijenik özellik: tanınma, adhezyon, reseptör Kolesterol (serbest kolesterol) Akışkanlık Lipoid matriks ile fiziksel etkileşim Membrana giriş-çıkış serbest Plazma membranında yüksek oranda! Ters kolesterol transportu Kolesterol lipid çift tabakada fosfolipid molekülleri arasına yerleşmiştir. Yapıdaki tek polar grup olan hidroksil grubu sulu ortama doğru yönelmiş ve fosfolipidlerin yüklü baş grupları ile etkileşimdedir.Polar olmayan steroid çekirdeği ve hidrokarbon zinciri ise fosfolipidlerin hidrokarbon kuyrukları arasında bulunur. Lipoid matriks ile fiziksel etkileşime girer. Plazma membranındaki kolesterolün tamamı serbest kolesteroldür. Kolesterol lipid çift tabakaya kovalent bağlarla tutunmadığı için, plazma membranından içeriye ve dışarıya serbestçe hareket edebilir. Bu özelliği nedeniyle kolesterol plazma lipoproteinlerine ya da lipoproteinlerden membrana aktarılabilmektedir. Kolesterol membran akışkanlığını düzenler; artırabilir ya da azaltabilir. Amfipatik lipidler Hidrofilik: gliserofosfolipidlerin polar başları(fosfat ve alkol grupları) Sfingofosfolipid(fosforilkolin) kolesterolün hidroksil grubu glikosfingolipidlerin şekerleri Lipofilik (hidrofobik): gliserofosfolipidlerde yağ asidlerinin hidrokarbon zincirleri Sfingomiyelinlerde,sfingozin+yağ asidi hidrokarbon zinciri kolesterolün steran halkası Amfipatik lipidler Hidrofilik: gliserofosfolipidlerin polar başları(fosfat ve alkol grupları) Sfingofosfolipid(fosforilkolin) kolesterolün hidroksil grubu glikosfingolipidlerin şekerleri Lipofilik (hidrofobik): gliserofosfolipidlerde yağ asidlerinin hidrokarbon zincirleri Sfingomiyelinlerde,sfingozin+yağ asidi hidrokarbon zinciri kolesterolün steran halkası Membran Proteinleri: Majör fonksiyonel moleküllerdir. Membran proteinleri, membranın spesifik fonksiyonlarından ve yapısal bütünlüğünden sorumludur Enzim D--hidroksibütirat dehidrogenaz / fosfatidil kolin Na+K+ATPaz, Ca2+ATPaz / kolesterol Kanal,taşıyıcı protein Yapısal komponent Antijen Reseptör Asetil kolin reseptörleri / kolesterol Plazma memb. >100 çeşit Sarkoplasmik ret. memb. <12 çeşit İntegral Proteinler-1 Lipid tabakalara gömülü/transmembran (alfa heliks yapı!) Bazı transmembran integral proteinler membranı bir kez kat eden tek bir hidrofobik bölge içerirken (LDL reseptörü, glikoforin), diğer bazıları çift tabakayı birden çok sayıda kateden birkaç hidrofobik bölgeye sahiptir (anyon kanal proteini, glukoz taşıyıcıları, G-proteine kenetli reseptörler). Fosfolipidlerle sıkı etkileşim ! Lipidlerle ilişki bozulursa denatürasyon ve aktivite kaybı ! Glisin, alanin, valin, lösin, izolösin, metiyonin ve fenil alanin, triptofan, prolin Deterjanlarla (SDS, deoksikolat, Triton X 100) veya organik çözücülerle izolasyon SDS-PAGE elektroforezi Eritrosit membranı anyon kanal proteini glikoforin : %60 karbohidrat içerir 16 oligosakkarid zinciri : 60-70 monosakkarid Myelin(beyin) lipofilin: proteolipid (hidrofobik lipoprotein) hidrofobik amino a. ve kovalan bağlı yağ a. >%65, organik çözücülerde çözünür, suda çözünmez Lipidlere kovalan bağlı proteinler Periferal proteinler olarak kabul edilirler. Protein palmitoil, miristoil, farnezil ve fosfatidil inozitol glikan (glikozilfosfatidil inozitol,GPI) gibi yapılarla membrana bağlıdır. Fosfolipaz C veya fosfolipaz D ile membrandan uzaklaştırılabilirler. Membranın Karbohidratları Glikoproteinler, glikolipidler Glukoz, galaktoz, mannoz, fukoz, N-asetil galaktozamin, N-asetil glukozamin, sialik asid* Proteinin katlanması, hücre yüzeyine transportu , reseptör fonksiyonları Hücre yüzeyinin antijenik özelliği Hücrelerin birbirlerini tanımaları ve adhezyon *(-) Yük nonspesifik agregasyonu engeller ! * Asialoglikoprotein varlığı (sialidaz) eritrositin yaşlı olduğunu gösterir ! Akışkanlık Lipid çift tabaka yapısı kararlı olmakla birlikte lipid ve proteinlerin hareket edebileceği kadar akışkandır.. Her bir yağ asidi hidrokarbon zincirinin –C–C− bağı çevresinde dönmesi ile hidrokarbon zincirleri sürekli hareket içindedir; bu hareket membrana akışkanlık kazandırır. Hidrokarbon zincirlerinin lipid çift tabaka içindeki hareket derecesi membran akışkanlığı olarak adlandırılır. Akışkanlığın derecesi temperatüre ve membranın lipid bileşimine bağlıdır Lipid bileşimi (Diyet !) yağ a. hidrokarbon zinciri C sayısı akışkanlık doymamışlık derecesi akışkanlık cis konfigürasyonu akışkanlık kolesterol: modülatör doymamış yağ a. arasında: akışkanlık doymuş yağ a. arasında: akışkanlık temperatür jel-kristallinlikid-kristallin: transisyon temp(Tm) vücut ısısında: likid zeminde jelöz bölgeler kolesterol, sfingomyelin, doymuş yağ asidi : jel aynı membranda özellikleri farklı olan bölgeler Permeabilite, transport ve reseptörler için akışkanlık ! akışkanlık permeabilite gap junction ve tight junction’lar: akışkanlık Düşük sıcaklıklarda hidrokarbon zincirleri düzgün sıralı halde ve hareketleri kısıtlı olduğundan lipid çift tabaka hemen hemen katı, jel-parakristalin durumdadır. Belirli bir sıcaklığın üzerinde ise hidrokarbon zincirlerinin termal hareket nedeni ile düzenleri bozulmakta ve membran sıvı halde bulunmaktadır. Membran lipid çift tabakasının katı halden sıvı hale geçtiği sıcaklık (transisyon temperatürü,Tm) olarak tanımlanır Biyolojik membranların heterojen bileşimlerinden dolayı Tm değerleri değişkendir. Tm, yağ asidi hidrokarbon zincirinin uzunluğuna ve doymamışlık derecesine ve membranın kolesterol içeriğine bağlı olarak değişmektedir. Doymamış yağ asidi oranı arttıkça, membranın Tm değeri düşmekte, dolayısıyla membranın akışkanlığı artmaktadır. Buna karşılık doymuş yağ asidi oranı arttıkça membranın Tm değeri artmakta ve akışkanlık azalmaktadır. Fosfolipid yağ açil zincirlerinin uzunluğu da Tm değerini etkilemektedir; yağ açil zincir uzunluğunun artması ile akışkanlık azalmaktadır. Doymuş ve doymamış yağ asitleri içeriği yanı sıra Tm’ nin diğer önemli bir belirleyicisi membranın kolesterol içeriğidir. Kolesterol membran akışkanlığını düzenleyen önemli bir modülatördür; akışkanlığı artırabilir veya azaltabilir. Yağ açil hidrokarbon zincirleri arasına girmiş olan katı düzlemsel halkalı yapıdaki steroid çekirdeğin akışkanlık üzerine iki etkisi vardır.. Membranın katı, jel-parakristalin halinde olması durumunda (Tm değerinin altında) kolesterol iki komşu fosfolipid yağ açil zincirleri arasına girerek van der Waals çekim etkisini azaltmakta ve zincirlerin düzenli bir şekilde paketlenmesini engelleyerek membran akışkanlığını artırmaktadır. Membranın sıvı-kristalin düzende olması durumunda (Tm değerinin üzerinde) kolesterolün katı, halka sistemi komşu yağ açil zincirlerinin C-C bağı çevresindeki dönme hareketlerini azaltmakta ve bu şekilde çift tabakanın merkezinde akışkanlığı azaltmaktadır Bu nedenle kolesterol membranın aşırı katı veya akışkan olmasını ılımlı hale getirmeye çalışan ve böylece akışkanlığı artırıp azaltabilen önemli bir modülatördür Ancak yüksek konsantrasyonlarda kolesterol varlığı membran akışkanlığını azaltmaktadır Membran asimetrisi İç-dış (transvers) asimetri Fosfolipidler : kolin içeren fosfolipidler: dış aminofosfolipidler: iç lateral mobilite yağ asidlerinin hidrokarbon zincirlerinin C-C bağları etrafındaki rotasyonel mobilite transvers mobilite (flip-flop) yavaş ! Proteinler Karbohidratlar (glikosfingolipidler ve glikoproteinler) Fosfolipid asimetrisini sağlayan taşıyıcılar Flippaz: aminogliserolipidlerin (PS, PE) iç tabakaya transportu, ATP bağımlı Floppaz: nonspesifik, dış tabakaya transport Skramblaz: nonspesifik, hücre içi [Ca2+] artışı ile aktive olur Pıhtılaşma (ekstrinsik yol) Hasarlı hücrelerin RES tarafından alınmaları Bölgesel asimetri gap junction ve tight junction protein-protein etkileşimi: protein matriks sinapslar mukoza hücrelerinde villuslar Fonksiyonel asimetri Na+K +ATPaz TEŞEKKÜRLER...