Replikasyon
advertisement

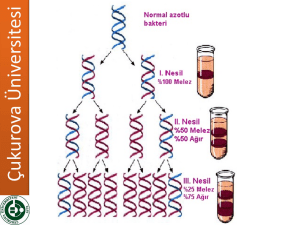
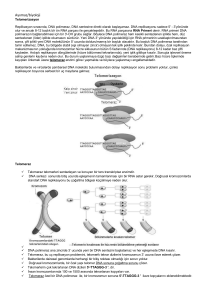
DNA REPLIKASYONU Bölüm 12 DNA Kendi kopyasını yapabilir, Tamir edilebilir, Rekombinasyon geçirebilir. Alternatif DNA replikasyon modelleri Watson & Crick azotlu bazlarin yerleşimi ve doğasından dolayı DNA ikili sarmalının her bir zincirinin tamamlayıcı zincir sentezi için kalıp oluşturabileceğini tahmin etmiştir. Fig. 12.1 1958: Matthew Meselson & Frank Stahl’s Experiment Semiconservative model of DNA replication (Fig. 3.2) Yarı saklı replikasyon için beklenen sonuçlar; Fig. 12.4 Equilibrium density gradient centrifugation (Box 3.1) I 1955: Arthur Kornberg E. Coli ile çalıştı DNA sentez mekanizmasını keşfetti İhtiyaç duyulan 4 ana bileşen 1. dNTP’ler: dATP, dTTP, dGTP, dCTP (deoxyribonucleoside 5’-triphosphates) (şeker-baz + 3 fosfat) 2. DNA kalıbı 3. DNA polimerazI (Kornberg enzimi olarak da bilinir) (DNA polimeraz II & III daha sonra bulundu) 4. Mg 2+ (DNA polimeraz aktivitesi için gerekli) 1959: Arthur Kornberg (Stanford University) & Severo Ochoa (NYU) DNA sentezinin 3 ana basamağı: 1. DNA polimeraz I, deoksiribozun 3’-OH grubu ile dNTP’nin 5’fosfat grubu arasındaki fosfodiester bağının oluşumunu katalizler. Bu reaksiyon için gerekli olan enerji NTP + (NMP)n (NMP)n+1 + P~P reaksiyonu ile karşılanır. 2. Zincir uzaması DNA polymeraz I’in komplementer dNTP’ları yapıya eklemesi ile sürer. • • rate ≤ 800 dNTPs/second Düşük hata oranı 3. Sentez yönü 5’ 3’ TANIMLAR Replikasyon orijini: Replikasyonun başlangıç noktası Replikasyon çatalı: Kromozom üzerinde replikasyonun olduğu noktada sarmala ait zincirlerin açılmasıyla ortaya çıkan bölgedir. Önce sentezin orijin noktasında meydana gelir ve replikasyon devam ettikçe ilerler.Replikasyon çift yönlü ise, orijinden itibaren zıt yöne doğru ilerleyen iki replikasyon çatalı oluşur. Replikon: bir orijinden bir replikasyon başladıktan sonra replike olan DNA’nın uzunluğunun bir birim olduğunu belirtmek için kullanılan terimdir. Replikasyon orijini (prokaryotik): Çift zincirli DNA molekülünün denatüre olarak açılması ve bazların serbest kalması ile başlar. replication bubble formunun oluşmasını sağlayan bu işlemden sonra replikasyon iki yönlü olarak devam eder. ~245 bp in E. coli REPLIKASYON ORIJINI (PROKARYOTIK): E. coli’de replikasyon 245 bç uzunluğunda olan ve oriC olarak adlandırılan tek bir orijinden başlamaktadır. Bakteri ve bakteriyofajlarda replikasyon tek bir noktadan başladığından kromozomun tümü replikondur. Replikasyon oriC’den heri iki yöne doğru hareket eder. Replikasyon ilerledikçe ayrı yönlere doğru birbirlerinden uzaklaşan iki replikasyon çatalı oluşur. Çatallar tüm kromozom replike olduktan sonra ter olarak adlandırılan sonlanma bölgesinde birbiriyle birleşir. Replikasyon orijini (ökaryotik): ÇOKLU REPLIKASYON ORIJINI (ÖKARYOTLARDA) 1. Ökaryotlarda birden çok replikasyon orijini (yeast 250-400, memelilerde 25,000’den fazla replikon) bulunur. * Daha fazla DNA içerirler (maya e.coli’den 4 kat drosophiladan ise 100 kat fazla DNA içerir) *Ökaryotik DNA pol’ın sn’de 50 nt olan sentez hızı bakteriyal polimerazlardan 20 kat daha yavaştır. ARSs (autonymously replicating sequences-özel replike olan diziler): 11 bç. Uzunluğundaki maya replikasyon orijinidir. Sentezin etkin başlamasını sağlayan kısa dizileri içerir. 20-80 replikon kümesi sıra ile aktive olur. Polimeraz çok büyük DNA dizilimi içinde ARS dizilerini, hücre döngüsünün S fazında başlayan bir mekanizma ile bulur. Hücre döngüsünün G1 fazında tüm ARS dizilerine bazı proteinler bağlanır (mayada 6 tane) ve orijin tanıma kompleksi(Origin Recognition Complex, ORC) oluşur. Kinazlar sentez aşamasında fosforilasyonda görev alan enzimlerdir. ORC’ye bağlanırlar ve DNA pol bağlanması için ön replikasyon kompleksi (pre-RC) oluştururlar.Kinazlar aktive olduklarında DNA sentezini tetiklerler ve her replikonda DNA sentezi tamamlanana kadar tekrar pre_RC oluşumunu engelleyerek kopyalanan ve kopyalanmamış DNA’ları birbirinden ayırırlar. Hücre döngüsü sırasında oluşan ve ökaryotlarda çok sayıda replikasyon orijini oluşumunu kontrol eden olaylar Fig. 12.16 DNA POLIMERAZ ENZIMLERI BAKTERILERDE DNA SENTEZINDE 3 POLIMERAZ VE DIĞER ENZIMLER GÖREV ALIR 1957, Kornberg ve ark. E. coli’den in vitro olarak DNA sentezini yönlendiren bir enzim saflaştırmış ve DNA polimeraz I olarak isimlendirmiştir. İn vitro DNA sentezi için enzimin ihtiyaçları; 1. 4 tip nükleotid trifosfat (dATP, dCTP, dGTP, dTTP= dNTP) 2. DNA kalıbı • 4 dNTP’den herhangi biri eksik ise yada ortamda NTP’lar yerine nükleotitler ve nükleotid difosfatlar varsa sentez olmamaktadır. • Kalıp DNA olmazsa sentez çok büyük oranda azalmaktadır. Kornberg’in izole ettiği enzim ile yapılan sentez şu şekildedir: DNA polimeraz I’in katalizlediği kimyasal reaksiyonun Her basamağında dNTP’ler yapıya eklenir. Uzayan DNA zincirine her bir nükleotidin katılım şekli DNA pol I’in özgüllüğüne bağlıdır. Öncü dNTP’de d-ribozun 5’ karbonuna bağlı fosfat grubu, ekleneceği dribozun 3’-OH grubuna kovalent bağla bağlanır. Fig. 12.8 DNA ZINCIRININ UZAMASI Ancak DNA pol I’in biyolojik fonksiyonu Kornberg tarafından tam olarak aydınlatılamamıştı, çünkü 1. Enzimin in vitro sentez hızı in vivo ya göre çok yavaştı 2. Tek zincirli DNA’yı çift zincirli DNA’dan daha etkin biçimde kopyalamaktaydı 3. DNA’yı sentezlediği gibi aynı anda yıkabilmekteydi. Kornberg- “DNA pol I in viro ortamda biyolojik olarak aktif DNA sentezleyebiliyorsa, hücre içinde de DNA sentezinin başlıca katalizörü olmalıdır” 1967, Goulian, Kornberg, ve Sinsheimer showed that DNA pol I’in biyolojik olarak aktif DNA sentezi yaptığını göstermişlerdir. Fig. 12.9 DNA POL I’IN BIYOLOJIK ROLÜ HALEN ŞÜPHELI IDI 1969 Delucia ve Cairns- DNA pol I aktivitesine sahip olmayan mutant bir E. coli suşu bulmuştur. Mutasyon polA1 olarak isimlendirilmiştir. Ancak mutant suş onarım yeteneği bakımdan oldukça yetersizdir. Bu gözlemlerden sonra Sonuç: 1. E. coli de in vivo replikasyon için en az bir pol daha olmalı 2. DNA pol I in vivo ikincil bir işlevi olmalıdır DNA POL II VE III DNA pol ı aktivite içeren normal hücrelerden elde edilen özgün enzimlerdir. DNA pol I, II ve III kalıptan DNA sentezi başlatamaz ve ancak primer adı verilen, var olan bir DNA zincirini kalıp boyunca uzatabilirler. 3’5’ ekzonükleaz aktivitesi: enzimin polimerizasyonu tek yönde gerçekleştirme, bir an duraksayıp, geri dönerek ilave edilen nt’leri çıkarabilme özelliğidir. 5’->3’ ekzonükleaz aktivitesi : DNA pol I’in özelliğidir. Enzim, sentezin başladığı uçtan itibaren nükleotitleri kesebilir ve sonra sentez yönünde işlemine devam edebilir. Bu nedenle DNApol I RNA primerini de uzaklaştırabilir. Ortamda pol III den daha fazla bulunur. Pol I primeri uzaklaştırır ve oluşan boşlukları doldurur. Pol II-> U.V hasarı nedeniyle oluşan mutasyonların tamirinde rol aldığı düşünülmektedir. Pol III -> polimerizasyondan aslı sorumlu olan enzimdir. Properties DNA Pol I Initiation of chain synthesis 5’-3’ polymerization + 3’-5’ exonuclease activity + 5’-3’ exonuclease activity + Molecules of polymerase/cell 400 DNA Pol II + + ? DNA Pol III + + 15 DNA POLIMERAZ I’IN KLENOW FRAGMANI DNA polimeraz I’in 5′ 3′ ekzonükleaz domain’i çıkarıldığı zaman geri kalan fragmana Klenow veya büyük fragman denir. Bu fragman polimerizasyon ve proofreading aktivitelerini korur. DNA POLIMERAZ I’IN 3′ 5′ EKZONUKLEAZ AKTIVITESI ILE HATALARI DÜZELTMESI DNA Pol III’ün holoenzim olarak adlandırılan aktif formu, iki takım, 10 farklı polipeptit zincirinden meydana gelmiş bir dimerdir. M. A 600.000 daltondur. Core (çekirdek) enzim holoenzimin polimerizasyon aktivitesi gösteren kısmıdır. g kompleks olarak adlandırılan ikinci bölgede 5 alt birim bulunur ve replikasyon çatalında enzimin kalıba oturtulmasında görev alır. b alt birimi, polimerizasyon sırasında enzimin kalıptan kopmamasını sağlar. P (pi), iki çekirdek replikasyon çatalında tutunmasını sağlar. polimerazın bir arad REPLİZOM Holoezim ve diğer proteinlerin replikasyon çatalında oluşturdukları büyük kompleks. E.coli polimeraz III’ün iki β altünitesi YUKARIDAN BAKIŞ Prokaryotik enzimler 3 çeşittir: Polymerase Polymerization (5’-3’) Exonuclease (3’-5’) Exonuclease (5’-3’) #Copies I Yes Yes Yes 400 II Yes Yes No ? III Yes Yes No 10-20 •Polimeraz I & III 5’->3’ yönünde replikasyon •Polimeraz II rolü tam olarak bilinmiyor •3’ -> 5’ exonükleaz aktivitesi= zincirin 3’ ucundan nükleotidlerin uzaklaştırılması •proofreading mekanizması olmadan hata oranı (mutasyon oranı) yaklaşık 1 x 10-6 •Proofreading ile hata oranı yaklaşık 1 x 10-9 6 farklı ökaryotik DNA polimeraz vardır DNA polimeraz α, δ (delta) ve ε ökaryotik çekirdek DNA’sı replikasyonu için gereklidir. DNA polimeraz b ve ζ (zeta) DNA tamirinde rol aldığı düşünülüyor. DNA polimeraz g (gama) mtDNA sentezinde rol alır. Prokaryotik/ökaryotik modeller (circular/linear kromozomlar) DNA REPLIKASYONU SIRASINDA BIRÇOK KARMAŞIK OLAY ÇÖZÜLMELIDIR DNA Sarmalı Açılmalıdır Replikasyon orijini (oriC) 9 ve 13 merden oluşan 245 bç lik bir bölgedir. DnaA ilk basamakta 9 mer’lik bu bölgeye bağlanarak sarmalın açılmasından sorumludur. •Bu bağlanma, sarmalın daha fazla açılmasında ve kararlılığında rol alan DnaB ve DnaC proteinlerinin bağlanmasını kolaylaştırır. •H bağlarını ATP enerjisi kullanarak kıran ve DNA’yı denatüre eden enzimlere HELİKAZ lar denir. SSBP’ler bu yapıyı tek zincire bağlanarak daha kararlı kılar. •Sarmal açıldıkça replikasyon çatalının önünde oluşan süpercoil yapı DNA gyrase (topoisomerase) tarafından gevşetilir. Fig. 12.10 Supercoiled DNA relaxed by gyrase & unwound by helicase + proteins: 5’ SSB Proteins Okazaki Fragments ATP 1 Polymerase III Lagging strand 2 3 Helicase + Initiator Proteins 3’ primase Polymerase III base pairs 5’ RNA primer replaced by polymerase I & gap is sealed by ligase RNA Primer 3’ Leading strand PRIMING DISCOVERY: TUNEKO OKAZAKI: 1985 1. RNA polimeraz nükleotid ekler, DNA polimeraz yapamaz 5’ T G ACA T T G C T C A 3’ A C UG U A A C G A G 5’ 2. DNA polimerazın primer ihtiyacı vardır DNA Sentezinin Başlaması İçin RNA Primerine Gereksinim Vardır •DNA pol III’ün polinükleotid zincirini uzatması için 3’ OH grubuna ihtiyacı vardır. •Kalıp DNA üzeirnde DNA’ya komplementer olan RNA parçası (5-15 nt) PRİMAZ enzimi •( RNA pol) tarafndan sentezlenir. •RNA primeri DNA pol I tarafından uzaklaştırılır ve zincir tamamlanır. •evrensel (virusler->prok->euks) Fig. 12.11 DNA SENTEZI 5′ 3′ YÖNÜNDE İLERLER Eğer zincir her zaman 5′ 3′ yönünde ilerlerse her iki zincir nasıl aynı anda sentez edilebilir? Bu soru 1960’larda Reiji Okazaki ve ark. tarafından çözüldü. Okazaki fragmanları, bir DNA zincirinde sentez edilen kısa DNA parçaları. Leading strand (öncül zincir): 5′ 3′ yönünde sürekli sentezin yapıldığı zincir Lagging strand (sonradan gelen zincir): Karşı zincirde 5′ 3′ yönündeki sentezin hücre tipine bağlı olarak birkaç yüz ile birkaç bin baz çiftli fragmanlar halinde sentez edildiği zincir Antiparalel zincirlerde DNA sentezi kesintili ve kesintisiz olarak gerçekleşir. •Kesintisiz (leading) •Kesintili (lagging) •(Okazaki fragments) 1000-2000 nt uzunluğunda •DNA pol I RNA primerinin uzaklaştırılması ve nt tatmamlanmasını yapar. •DNA ligaz fragmentleri birleştirir. Fig. 12.12 DNA ligase seals the gaps between Okazaki fragments with a phosphodiester bond (Fig. 3.7) Sentez kesintili ve kesintisiz zincirlerde aynı anda yapılır Fig. 12.13 I PROOFREADING (HATA OKUMA) VE DÜZELTME DNA REPLIKASYONUNUN AYRILMAZ PARÇASIDIR Exonuclease proofreading (DNA polymerase I & III 3’->5’ ekzonükleaz aktivitesine sahiptir) DNA pol III’ün Epsilon altünitesinin işlev göremediği mutant E. coli suşlarında DNA sentezi sırasında hata hızı önemli ölçüde artar. Prokaryotlarda DNA Replikasyon Modeli Fig. 12.14 DNA REPLIKASYONUNDA BIRÇOK ENZIM VE PROTEIN FAKTÖR ROL OYNAR E. coli replikasyonunda, DNA polimeraz dışında 20’den fazla sayıda farklı enzim ve protein rol alır. Bu kompleksin tümünü DNA replikaz sistemi veya replizom denir. Replikasyonun başlaması için ikili sarmalı ayrılmasını sağlayan enzimler helikaz’lardır. Zincir ayrılması ile oluşan topolojik stress topoizomerazlar ile giderilir. Ayrılmış zincirler DNA bağlayan proteinler ile stabilize edilir. Primazlar, genellikle kısa RNA parçacıkları olan primerleri sentezler. RNA primeri çıktıktan sonra boşluk DNA ligaz ile kapatılır. ÖKARYOTIK DNA SENTEZI PROKARYOTIK DNA SENTEZINE BENZER ANCAK DAHA KARMAŞIKTIR Temel gereksinimleri benzerdir ancak ÇOKLU REPLIKASYON ORIJINI 1. Ökaryotlarda birden çok replikasyon orijini (yeast 250-400, memelilerde 25,000’den fazla replikon) bulunur. * Daha fazla DNA içerirler (maya e.coli’den 4 kat drosophiladan ise 100 kat fazla DNA içerir) *Ökaryotik DNA pol’ın sn’de 50 nt olan sentez hızı bakteriyal polimerazlardan 20 kat daha yavaştır. ARSs (autonymously replicating sequences-özel replike olan diziler): 11 bç. Uzunluğundaki maya replikasyon orijinidir. Sentezin etkin başlamasını sağlayan kısa dizileri içerir. 20-80 replikon kümesi sıra ile aktive olur. Polimeraz çok büyük DNA dizilimi içinde ARS dizilerini, hücre döngüsünün S fazında başlayan bir mekanizma ile bulur. Hücre döngüsünün G1 fazında tüm ARS dizilerine bazı proteinler bağlanır (mayada 6 tane) ve orijin tanıma kompleksi(Origin Recognition Complex, ORC) oluşur. Kinazlar sentez aşamasında fosforilasyonda görev alan enzimlerdir. ORC’ye bağlanırlar ve DNA pol bağlanması için ön replikasyon kompleksi (pre-RC) oluştururlar.Kinazlar aktive olduklarında DNA sentezini tetiklerler ve her replikonda DNA sentezi tamamlanana kadar tekrar pre_RC oluşumunu engelleyerek kopyalanan ve kopyalanmamış DNA’ları birbirinden ayırırlar. BIDIRECTIONAL REPLICATION Eukaryotes vs. prokaryotes Linear, multiple origins vs. circular, one origin Theta mode q •2 replication forks •In eukaryotes Hücre döngüsü sırasında oluşan ve ökaryotlarda çok sayıda replikasyon orijini oluşumunu kontrol eden olaylar Fig. 12.16 6 farklı ökaryotik DNA polimeraz vardır DNA polimeraz α, δ (delta) ve ε ökaryotik çekirdek DNA’sı replikasyonu için gereklidir. DNA polimeraz b ve ζ (zeta) DNA tamirinde rol aldığı düşünülüyor. DNA polimeraz g (gama) mtDNA sentezinde rol alır. ÖKARYOTIK REPLIKASYON Polimeraz alfa replikasyonun başlamasından sorumludur (50,000/hücre) Polimeraz alfanın iki alt ünitesi RNA primerini sentezleyen primaz olarak işlev görür. Sonra diğer alt birim primerin ucuna nt takarak sentezi sürdürür. Pol a’nın işlev kapasitesi düşüktür. Bu nedenle RNA primerine kısa bir DNA dizisi eklendikten sonra, polimeraz değişimi olarak billinen işlem gerçekleşir. Pol a kalıptan ayrılır yerine pol d (delta) geçer. Böylece sentez hızı 100 kat arta, ve 3’->5’ ekzonükleaz aktivitesi kazanılır. Pol e, pol d (delta) ile aynı özelliklere sahiptir ancak değişik hücre koşullarında çalıştığı düşünülür. Tüm işlemler kesintili ve kesintisiz zincir için aynıdır. Doğrusal kromozomların uçlarının replikasyonu so •Kesintili zincirde RNA primeri uzaklaştırıldığında kromozomun ucunda serbest 3’OH grubunu sağlayacak kalıp zincir olmadığı için sentez yapılamaz. •Kromozom RNA primerini boyu kadar kısalır. Çözüm: Telomer ve Telomeraz •Tetrahymena’da Telemorlerin çoğu 5’-TTGGG-3’ dizisi ile sonlanmaktadır. Telomeraz her replikasyonda telomerin kısalmasını önlemek için TTGGGG tekrar dizilerini kromozomun ucuna eklemektedir. İlave edilen diziler “saç tokası” gibi kıvrılır ve karşı karşıya gelen G’ler arasında H bağı kurulur ve serbest 3’-OH grubu elde edilir. DNA pol I boşluğu doldurur.Daha sonra saç tokası kırılır ve DNA kaybı engellenir. Fig. 3.19 Synthesis of telomeric DNA by telomerase Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.