moleküler biyolojide kullanılan yöntemler
advertisement

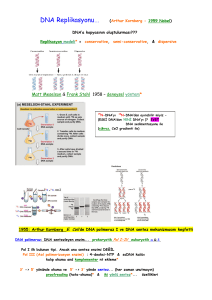
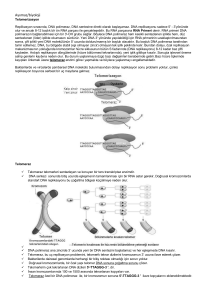
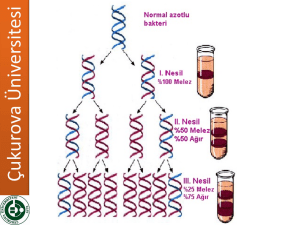
6.3.2016 Dersin içeriği MOLEKÜLER BİYOLOJİDE KULLANILAN YÖNTEMLER • Nükleik asitler • Hücre parçalama yöntemleri • Ayırma ve saflaştırma yöntemleri (filtrasyon, diyaliz, çöktürme santrifüjleme vb) • DNA izolasyonu ve analizi • RNA izolasyonu ve analizi • DNA’nın polimeraz zincir reaksiyonu ile çoğaltılması • Rekombinant DNA teknolojisi • Nükleik asit hibridizasyonu yöntemleri • Protein saflaştırma yöntemleri GİRİŞ • Ara Sınav 50 • Ödev 30 • Performans Görevi (Seminer) 20 Yıl İçinin Başarıya 55 Final 45 • Doğada milyonlarca farklı canlı türü bulunur. Bununla beraber canlılar hücre ve molekül düzeyinde araştırıldığında tek bir kalıp planı olduğu ortaya çıkar. • Buna göre moleküler biyolojinin temel amacı canlılarda hücre yapısını oluşturan yapısal ve işlevsel analizini yapmaktır. Hücreyi oluşturan temel organik moleküller başlıca iki grupta toplanabilir: • Bir hücrenin ağırlığının yaklaşık %99 u C, H, N ve O den oluşur. • İnorganik ve organik kimyasal bileşenlerden oluşan hücrelerde C atomlarından türevlenen organik bileşikler yaşam molekülleri olarak kabul edilirler. • Küçük moleküller: hücrenin temel kimyasal yapıtaşlarının sentezinde ve enerji elde edilmesinde kullanılırlar • Büyük moleküller (Makromoleküller): küçük moleküllerin polimerleri olan makromoleküller biyolojik süreçlerin özgüllüğünden ve biyolojik bilginin transferinden sorumludurlar. • Polimerik yapıdaki büyük moleküller monomerik yapıdaki küçük moleküllerin kovalent olarak birbirlerine bağlanmasıyla oluşurlar. 1 6.3.2016 Hücrelerde küçük organik moleküllere ait başlıca dört temel aile vardır: basit şekerler yağ asitleri aminoasitler nükleotitler • Makromolekülerin kendilerini oluşturan küçük moleküllerle çok az benzerlikleri vardır. • Makromoleküller arasında özellikle nükleik asitler ve proteinler hücrenin ve dolayısıyla bireyin yapısal oluşumu ve canlılık işlevleri açısından çok önemli rol oynarlar. • Nükleik asitler canlılık özelliklerini yönettikleri için biyolojik açıdan en önemli makromoleküllerdir. • Nükleik asitlerden DNA canlıların tümüne yakın kısmında, genetik bilginin ana deposudur. Bu bilgi RNA molekülleri biçiminde kopyalanır (transkripsiyon) ve daha sonra proteine çevrilir (translasyon). • Protein sentezi nükleotit dizisindeki şifreye göre yapılır. Moleküler biyolojide genetik bilginin akış yoluna ana kural (sentral dogma) denir. Moleküler biyolojinin santral dogması • Bir makromolekül küçük molekül ağırlıklı alt birimlerin birbirine eklenmesiyle oluşan uzun ve zincir biçiminde polimerdir. • Makromoleküller Proteinler polisakkaritler nükleik asitler • Nükleik asitlerin temel rolü genetik bilginin yaşam boyunca korunmasının nesilden nesile geçişinin sağlanmasıdır. • Genetik bilginin önemli bir kısmının anlatımı proteinler biçiminde ortaya çıkar. • Yani nükleik asitler bireyin oluşumunu ve türüne özgü karakterlerini kazanmasını proteinler aracılığıyla yönetirler. • Canlı bir hücrede pek çok metabolik olay ve reaksiyon proteinler tarafından yürütülür. • Proteinler yapısal eleman olarak da önem taşırlar Kan plazma proteinleri Hormonlar Antikorlar Enzimler Kloroplast Mitokondri Hücre duvarı ve proteinde yer alan proteinler vb yapısal ve işlevsel protein gruplarıdır Her hücre tipinde binlerce farklı tipte çeşitte protein bulunur 2 6.3.2016 Nükleik asitler • Bu nedenlerle moleküler biyolojiye ilişkin çalışmalarda özellikle nükleik asitlerin ve proteinlerin yapısal ve işlevsel analizlerinde moleküler biyolojik yöntemlerin bilinmesi ve kullanımı büyük önem taşır. Hücrenin canlılığının sürdürülmesi ve neslinin devam ettirilmesini sağlayan komponentler Hem ökaryotik hem de prokaryotik hücrelerde iki tip nükleik asit bulunur. DNA RNA Nükleik asitlerin yapısı Nükleik asitler 3 bileşenden oluşur - şeker - baz - fosfat (PO43- ) • Deoksiribonükleik asit Genetik bilgiyi (şifreyi) taşıyan ve bu bilginin kuşaklar boyunca aynı şekilde korunmasını sağlayan polimer. • 3 farklı RNA vardır (mRNA, rRNA, tRNA) • Herbiri farklı işleve sahiptir. DNA’nın taşıdığı genetik bilgiyi kullanarak hücre için gerekli proteinleri sentezlerler. Bazlar 1. Purin bazlari 2. Pirimidin bazlari Şekerler Deoksiriboz Riboz DNA RNA Pirimidin halkası Purin halkası Adenin (A) Guanin (G) Sitozin (C) Timin (T) Urasil (U) Riboz Deoksiriboz RNA DNA DNA RNA Nükleozitler Şeker+ Baz = nükleozit A,G,C,T A,G,C,U Nükleotitler Şeker + baz + fosfat = nükleotit Riboz veya deoksiribozun 1. C atomuna purin veya pirimidin bazları glikozidik bağlarla bağlanır Nükleotitlerin birleşmesi ile de polinükleotitler oluşur. 3 6.3.2016 Polinükleotit (nükleik asit) • Zincir sentezlenirken her bir deoksiribozun 3’ karbonuna diğer bir deoksiribozun 5’ucundaki fosfat grubu ile forfodiester bağı ile bağlanır • Nükleik asitlerde, birbirini izleyen şeker ve fosfat molekülleri nükleik asitlerin iskeletini oluşturur. (fosfodiester bağı: tek bir fosfat ester bağı ile iki ayrı şekere bağlanmıştır) Zincirdeki ilk nükleotit 5’ pozisyonunda daima PO4 grubu içerir. Diğer ucunda ise zincire son eklenen deoksiribozun 3’ pozisyonunda ise serbest bir OH grubu bulunur. • Tek zincir halindeki nükleik asitler kural olarak daima 5’ ucu solda ve 3’ ucu sağda olacak şekilde 5’ 3’ yönünde yazılır Örn 5’-AAGCTCTTAGG-3’ • RNA’nın yapısı da DNA gibidir RNA molekülleri ribonükleotit ünitelerinin 5’ 3’ yönünde fosfat diester köprüleri kurmasıyla oluşmuştur. DNA zincirinin omurgasında bulunan bütün fosfat grupları tamamen iyonize durumdadır ve negatif yüklüdür. BU NEDENLE DNA BİR ASİTTİR RNA’nın DNA’dan farkları • RNA yapısındaki ribozun 2’ OH grubundan dolayı DNA’dan daha az stabildir Alkalilerle muamele edildiğinde kolaylıkla 2’ - 3’ siklik monofosfat türevlerine parçalanır. • RNA’nın yapısındaki şeker ribozdur • Primidin bazlarından timinin (T) yerine urasil (U) geçmiştir. • Çoğunlukla tek zincir halindedir, ancak kendi etrafında geriye dönebilir,baz çiftleri oluşturarak molekülün özel bir konformasyon kazanmasını sağlayabilir • Alkalilere dirençli değildir. Genelde DNA’ya oranla çok daha kolay nükleotitlerine parçalanır. 4 6.3.2016 DNA’nın çift zincirli yapısı • RNA’nın hücrede 3 kritik rolü vardır • Elçi RNA (mRNA): DNA’nın bir zincirindeki genetik bilgiye komplementer olan bilgiyi içerir. • Transfer RNA (tRNA): protein sentezindeki adaptör moleküllerdir. Nükleotit dilindeki genetik bilgiyi aminoasit diline dönüştürür. • Ribozomal RNA (rRNA): hücrenin protein sentez sistemi olan yapısal ve katalitik bileşenidir. • DNA’nın çift polinükleotit zincirinden oluştuğunu Watson ve Crick (1953) ispatlamışlardır. • • Hücre içinde DNA çift zincirli formda bulunur. Her kromozom iki zincirli bir DNA taşır Bu zincirlerin her biri fosfodiester bağlarıyla bağlı yüzbinlerce ya da milyonlarca nükleotit içerir. • Şeker ve fosfat DNA’nın omurgasını oluşturur • Bazlar ise diğer zincirdeki bazlarla eşleşir. Bütün DNA moleküllerinde Adenin, Timin ile çift H bağı oluşturur Guanin ise Sitozin ile 3’lü Hidrojen bağı oluşturur. A = T Guanin • Adenin ile Timine • Guanin ile de Sitozin’e birbirinin komplementeri denir Çift zincirli bir DNA molekülündeki iki zincir de birbirinin komplementeridir. İki iplik birbirinin komplementeri olduğu için DNA’daki tüm bilgi her iki iplikte de kopyalanmış durumdadır. Bu durum DNA’nın kopyalanmasındaki temel özelliktir =1 C Sitozin Adenin Timin DNA molekülündeki Adenin miktarı Timin miktarına; Guanin miktarı da Sitozin miktarına eşittir. • DNA molekülündeki A+T miktarının G+C miktarına oranı hiçbir zaman 1 olmaz. Daima 1’de büyük veya küçüktür. • Bu oran aynı tür içindeki canlılar arasında değişmez, türden türe ise farklılıklar gösterir • Çift sarmaldaki polinükleotitler antiparalel şekilde oluşur. • Bir polinükletit 5’ 3’ yönünde; karşındaki polinükleotit ise 3’ 5’ yönünde ilerler H bağları kovalent bağlardan daha zayıf olduğu için iki DNA ipliği çeşitli koşullarda kolayca ayrılıp tekrar birleşebilirler G DNA’nin çift sarmal yapısı • • İki zincir birbirinin etrafında sarılarak ikili sarmal oluşturur. DNA’nın 3 boyutlu bilinen 3 farklı yapısı vardır: A-DNA (Sağ el formu) B-DNA (sağ el formu) Z-DNA (sol el formu) Hücrelerde B-DNA hakimdir A-DNA su kaybetmiş DNA örneklerinde görülebilir Z-DNA DNA’nın metilasyona uğramış kısımları bu formda görülebilir. Sağ el Sağ el Sol el 5 6.3.2016 B-DNA’nın çift sarmal yapısı • Watson ve Crick tarafından belirlenen ve hücrelerde hakim olan yapıdır. İkili sarmal halindeki DNA birbirinden ayırt edilebilen büyük oluk ve küçük oluk adında iki oluktan oluşur. Herbir baz çifti arasındaki uzunluk 0.34 nm Büyük oluk Küçük oluk Sarmalın her döngüsü yaklaşık 10 baz çifti (bp) içerir (3.4 nm). 1 kb DNA 100 sarmal döngü içerir ve 0.34 µm uzunluktadır. DNA molekülünün büyüklüğü • Bir DNA molekülünün büyüklüğü, herbir molekülde binlerce nükleotit bazları veya baz çiftleri şeklinde ifade edilir. • İnsan genomunda yaklaşık 3 milyar nükleotit bulunmaktadır. • E. coli’de ise 4 milyon 640 bin nükleotit vardır. • 1000 bazlık bir DNA molekülü 1 kilobaz içerir (kb) • Eğer DNA ikili sarmal ise 1 kilo baz çifti (kbp) denir. DNA’nın denatürasyonu DNA’nın süpersarmal yapısı • Gevşek bir DNA molekülünün baz çifti sayısı bilindiğinde heliks döngü sayısı tahmin edilebilir. • Ancak, E. Coli kromozomu lineer hale getirilirse 1 mm’den daha uzun olacaktır. • Bu uzunluk E.coli’nin kendi çevresinden 400 kat daha fazladır. • Böyle küçük bir alan içerisine bu kadar fazla DNA nasıl paketlenir? DNA’nın replikasyonunda ve protein sentezinin ilk evresi olan transkripsiyonda sentezin başlayabilmesi için iki DNA ipliğinin birbirinden ayrılması gerekir. Bazların H bağları kırılarak birbirinden çözülür ve ayrılır. Hücrede (in vivo) doğal olarak gerçekleşen çözülmesi ayrılması ve tekrar birleşerek eski formunu kazanması deneysel olarak hücre dışında da (in vitro) gerçekleştirilebilir. • Bu, DNA’nın süpersarmal adlı bir işlemle kıvrılarak katlanmasıyla gerçekleşir. DNA hücrelerden izole edildiği oda sıcaklığına yakın ortamda fizyolojik tuz konsantrasyonunda muhafaza edildiğinde ikili sarmal değişmeden kalır. A=T G≡C Eğer sıcaklık arttırılırsa iki zincir arasındaki H bağları kırılarak birbirinden ayrılır. Bu işleme denatürasyon (erime) denir. Tek zincirli ve çift zincirli nükleik asitler 260 nm’de UV ışığı absorblamada farklılık gösterdiklerinden bu durum deneysel olarak belirlenebilir. Burada esas nükleik asit bazlarının UV ışığı absorblamalarıdır. Çift zincirli DNA’ nın absorbsiyonu tek zincire göre oldukça azdır G ve C eşleşmesi daha güçlüdür. DNA’daki GC içeriği arttıkça Tm değeri de artar. Renatürasyon: Sıcaklık artışıyla DNA çözülmeye başladığında absorbansda ani artış gözlenir. DNA içeren çözeltide sıcaklık yükseltilerek alınan en yüksek absorbans değerinin yarısı için gerekli sıcaklık derecesine Tm (erime sıcaklığı-melting temperature) denir. Isıtılarak denatüre edilen DNA yavaşça soğutulursa DNA’nın doğal yapısı olan çift zincir yeniden oluşur. Bu olaya da renatürasyon denir. Denatürasyon ve renatürasyon DNA hibridizasyon tekniğinin esasını oluşturur. Hibridizasyon: iki farklı DNA önce denatürasyonla ayrılır sonra ayrılan farklı zincirlerin eşleşmeleri sağlanır. Tm: yarı maksimal absorbans için gerekli sıcaklık 6 6.3.2016 DNA Replikasyonu (DNA çoğalması, DNA ikileşmesi, DNA sentezi) • Bir hücrenin bölünebilmesi için DNA’nın da çoğalması gerekir. • DNA’nın replikasyonu • DNA replikasyon mekanizmasının anahtarı, DNA ikili sarmalındaki iki ipliğin de aynı bilgiyi taşımasıdır (birbirleriyle komplementer olmaları). • Böylece oluşan iki yavru hücre de aynı DNA’ya sahip olur. 37 • Bu yüzden DNA’nın replikasyonu yarı korunumludur (semikonservatif). Yani oluşan ikili sarmalin zincirlerinden biri yeni sentezlenen zincirken, diğeri eski var olan zincirdir. • Replikasyon sırasında iki atasal iplik ayrılır, tamamlayıcı yavru ipliklerin oluşmasında kalıp (template) olarak görev yapar. • Daha sonra oluşan yeni çift zincirli DNA moleküllerin bölünme sırasında iki yavru hücreye geçer. Atasal (eski) iplik Yavru (Yeni) iplik Yavru (Yeni) iplik 39 Replikasyonun aşamaları 1) replikasyon orijinine özel başlangıç proteinlerinin bağlanması 40 • Prokaryotlarda tek bir replikasyon orijini vardır. Ökaryotlarda çok fazla orijin bulunmaktadır • • Replikon: tek bir orijinden başlayıp sentezlenen DNA’ya replikon denir (insan genomumda 30.000 orijin bulunmaktadır) • Mayalarda 250-400 arasındadır.. Replikasyon orijini: replikasyonun başladığı genomdaki sabit bir noktadır. Prokaryotlarda OriC (E.coli) olarak bilinir. Replikasyon orijini özel başlangıç proteinleri (dnaA) tarafından tanınan ve yaklaşık 300 bazlık özel bir diziden oluşur.(E.coli’de 245 bazdır) . Replikasyon orijini AT zengin bölgelerdir. Böylece zincirlerin ayrılması daha kolay olur. 41 42 7 6.3.2016 2) Çift zincirin açılması • • Replikasyonun ilk aşamasında Orijin bağlama proteinleri (dnaA) orijine bağlanır ve kompleksi açmak için erimeyi gerçekleştirir. • Bu bağlanma daha sonra zincirin açılmasını kolaylaştırır. dnaA orijine bağlandıktan sonra helikaz enzimi (dnaB) çift sarmaldaki H bağlarını kırarak zinciri açar ve replikasyon çatalı oluşur. Replikasyon çatalı: İpliklerin ayrıldığı ve yeni DNA sentezinin gerçekleştiği noktaya replikasyon çatalı denir. Replikasyon çatalı 43 • Genellikle orijinden itibaren iki replikasyon çatalı çift yönlü olarak çalışır ve iplikler terminale ulaşıncaya kadar ayrı ayrı kopyalanır. • DNA replikasyonu E.coli hücrelerinde açığa kavuşturulduğundan, Replikasyon için E. Coli örnek oluşturmaktadır. • Ökaryotlarda replikasyon çok daha karmaşık olmakla birlikte temelde bakteriyel DNA replikasyonuna benzer. 44 • Replikasyon süresince helikaz DNA üzerinde ilerleyerek sürekli olarak ikili zinciri açar. • Helikaz sarmalın çözülmesini çok hızlı gerçekleştirir. • Helikaz ile çift zincir açıldıktan sonra, tek iplikler üzerine tek iplik bağlama proteinleri (ssb) bağlanarak ipliklerin yeniden birleşmesini (renatürasyonunu) önlerler. Zincir açıldıkça oluşan süpersarmal yapı topoizomerazlar (DNA giraz)tarafından giderilir. 45 3) RNA primer sentezi 46 • Yeni DNA sentezinde nükleotitlerin eklenmesini (zincirin uzamasını) sağlayan enzim DNA polimerazdır. • Ancak hiçbir DNA polimeraz DNA sentezini başlatamaz. Bu enzim sadece daha önce var olan 3’-OH grubuna nükleotit ekleyebilir. • Bütün bilinen DNA polimerazlar deoksirobozun 3’ OH grubuna nükleotit ekleyebilirler. • Bu nedenle yeni DNA zincirinin başlayabilmesi için mutlaka önce primer sentezlenmesi gerekir. • Bu nedenle DNA’nın replikasyonu daima 5’-3’ yönünde olur. • 47 Primer: 10-15 nükleotit dizisinden oluşmuş kısa bir RNA parçacığıdır ve özel bir RNA polimeraz olan primaz enzimi tarafından sentezlenir. Daha sonra zincirin uzaması DNA polimeraz tarafından gerçekleştirilir. 48 8 6.3.2016 4) Zincirin uzaması • Helikaz enzimi ile çift zincir açılır açılmaz primaz enzimi replikasyon çatalının hemen başında yerini alır ve gerektikçe primer sentezler. • Replikasyon çatalında yer alan helikaz ve primaz enzimlerinin ardından açılan her iki polinükleotit kalıbına yerleşen DNA polimeraz enzimi sentezi her iki zincirde de sürdürür. • Primer eklendikten sonraDNA polimeraz sentezi sürdürür. • DNA polimeraz yeni DNA zincirini yalnızca 5’3’ yönünde gerçekleştirir. • Ancak, kalıp DNA zincirlerinin antiparaleldir (zincirin biri 5’-3’, diğeri 3’-5’ yönünde) • Bu durumda DNA polimeraz ters zincirdeki sentezi diğeri ile aynı anda nasıl gerçekleştirir? 49 50 • 3’-5’ yönündeki sentez mekanizması Okazaki tarafından çözülmüştür • Okazaki DNA sentezinde bir zincirdeki sentezin 5’-3’ yönünde sürekli devam ettiğini karşı zincirde ise kesikli sentezin gerçekleştiğini belirlemiştir. • Açılan kalıp DNA zincirlerinde sadece birinde komplementer polinükleotit sentezi sürekli olur. Her zaman yeni bir nükleotitin ekleneceği serbest bir 3’-OH grubu olduğundan kesiksiz olarak DNA sentezi gerçekleşir. • DNA sentezinin kesintisiz sürdüğü zincire kesintisiz DNA zinciri (leading strand), diğerine ise kesintili DNA zinciri (lagging strand) denir. • Kesikli zincirde zincir uzaması replikasyon çatalına ters yönde olur. • Kesintisiz DNA zinciri tek bir primer kullanarak ve başlangıç noktasından başlayarak 5’-3’ yönünde replikasyon çatalının açılma yönünde sentezi sürdürür. • Kesintili zincirde ters yönde herbiri ayrı primer gerektiren 1000-2000 nükleotitlik fragmanlar sentezlenir ve bunlar birbirine eklenir. Bu nükleotit fragmanlarına Okazaki fragmanları denir. • Diğer zincirde ise ters yönde ve kesikli ve kısa zincirler halinde sentez gerçekleşir. 51 • Kesikli zincirde primaz tarafından defalarca RNA primerleri sentezlenerek serbest 3’-OH grubunun oluşmasını sağlarlar. 52 DNA polimerazlar: Nükleotitlerin eklenmesini katalizleyen enzim. Bilinen bütün DNA polimerazlar DNA’yı 5’-3’yönünde sentezler. Zincirdeki her yeni nükleotitin öncüsü bir deoksiribonükleotit trifosfatdır (dNTP) dNTP: dATP, dTTP, dGTP, dCTP, • Bunun aksine kesiksiz zincire sadece başlangıç yerinde olmak üzere bir defa primer eklenir. dNTP’lerin iki terminal fosfatı uzaklaştırılır, ve internal fosfat büyüyen zincirin deoksibozunun 3’OH grubuna bağlanır. 53 54 9 6.3.2016 • E. Coli’de 3 tip DNA polimeraz belirlenmiştir. • DNA polimeraz II (Pol ll): • DNA polimeraz I (Pol I): Birden fazla aktiviteye sahiptir. RNA primerlerini uzaklaştırıcı 5’-3’ ekzonükleaz aktivitesi (bu özellik sadece bu enzimde vardır) DNA sentezi kesintili zincirde kalan boşlukları tamamlar. Daha önce oluşmuş olan RNA primerlerini uzaklaştırır ve yeni sentez yapar. Hücredeki konsantrasyonu DNA pol lll’den daha fazla DNA pol l: 400 mol/hücre DNA pol lll: 10-20 mol/hücre görevi henüz olarak bilinmiyor. Ancak çeşitli çevresel faktörlerle (UV ışık, kimyasallar vs) oluşan DNA tamirinde kullanıldığı düşünülüyor. • Bu onarımlar DNA replikasyonu tamamlandıktan sonra yapılır. • DNA polimeraz III (Pol lll): Her iki zincirde de replikasyonu gerçekleştiren ana enzimdir. Polimerazlar Kofaktör olarak Mg iyonlarına gereksinim duyarlar 55 • Ayrıca her 3 DNA polimerazın da 3’- 5’ ekzonükleaz aktivitesi vardır. • Polimerizasyon sırasında yanlış bir nükleotitin zincire eklenmesi durumunda, kalıp DNA’da bu bazın tamamlayıcısı olmayacaktır. Bu hatanın düzeltilmesi için geriye dönüp bu nükleotitin çıkarılması ve ve yerine doğrusunun bağlanması gerekir. 56 • Kesikli zincirde Pol lll daha önce sentezlenen DNA’ya ulaşıncaya kadar sentez yapar. Bu noktada Pol lll durur. • • Her 3 polimeraz da 3’- 5’ ekzonükleaz aktiviteleri ile bu düzeltmeyi (proofreading) yapabilmektedirler . Sentezde rol alan bir sonraki enzim olan DNA polimeraz l, önce, RNA primerlerini uzaklaştırır ve bunların yerini DNA ile doldurur. • Son fosfodiester bağı ise DNA ligaz ile yapılır. • Hata düzeltme fonksiyonu sayesinde replikasyondaki hata oranı milyarda 1’dir 57 58 5) Terminasyon ve ayrılma • DNA ligaz bazların 5’ fosfat uçları ile 3’OH uçlarını birleştirirek ayrı ayrı oluşan DNA parçalarını fosfodiester bağları ile birleştirir. • DNA ligaz DNA parçalarını birleştirebilen tek enzimdir. • İki replikasyon çatalı oriC’nin yaklaşık 180° karşısındaki bölgede buluşur. Bu bölgede Helikazı durduran tus gen ürününün bağlandığı bir sonlanma noktası vardır. • Replikasyon tamamlandığında iki halka birbirine bağlı olarak kalmaktadır. • Bunlar bir topoizomeraz olan topoizomeraz IV aracılığıyla birbirinden ayrılır ve daha sonra membran tutunma yerlerinin bir kısmıyla hareket ederek iki yavru hücreye ayrılırlar 59 60 10 6.3.2016 Özet ÖZET • • DNA replikasyonundaki enzimler DNA’nın replikasyonunda çok sayıda protein yer alır. Bunların hepsine DNA replikaz sistemi veya replizom denir Orijin bağlama proteinleri: replikasyon orijinine bağlanarak ilk denatürasyonu gerçekleştirir. Helikaz :Replikasyonun başlaması için ikili sarmalı ayrılmasını sağlayan enzimler Topoizomerazlar: Zincir ayrılması ile oluşan kıvrımları giderir. Tek zincir bağlama proteinleri: Ayrılmış zincirler tek zincir bağlama proteinleri (ssb) ile stabilize edilir. Primazlar, RNA parçacıkları olan primerleri sentezler. DNA polimeraz III 5’-3’ OH yönünde dNTP’leri ekleyerek sentezi gerçekleştirir. DNA polimeraz l primerleri çıkararak boşlukları yeni DNA ile doldurur DNA ligaz: RNA primeri çıkarılıp yeni DNA sentezi yapıldıktan sonra boşluğu kapatır. 61 • • • • • Replikasyon orijinine orijin bağlama proteinleri bağlarak bu bölgeyi denatüre eder. Helikaz enzimi zinciri iki yönlü olarak açarak replikasyon çatalları oluşturur. Zincir bağlama proteinleri her iki zincire de tutunarak zincirin tekrar birleşmesini önler DNA primaz enzimi her iki zincir üzerinde de primerleri sentezler. DNA pol lll 5’-3’ yönünde dNTPleri zincire ekleyerek sentezi gerçekleştirir. Ters yönde olan zincirde ise replikasyonda ters yönde gerçekleşir. Burada primer 1000-1500 bazda bir yeni primer sentezlenerek Pol lll Okazaki fragmentlerinin sentezini yapar. DNA Pol I sentezlenen primerleri zincirden uzaklaştırır ve yerine DNA sentezler. Son nükleotitler arasındaki boşluk DNA ligaz ile kapatılır. 62 11