MOLEKÜLER BİYOLOJİ
(2011-2012)
Doç. Dr. Ercan ARICAN
İ.Ü. Fen Fakültesi
Moleküler Biyoloji ve Genetik Bölümü
A. Hücreler ve Makromoleküller
A1. HÜCRESEL SINIFLANDIRMA
A. Hücreler ve Makromoleküller
A1. HÜCRESEL SINIFLANDIRMA
Öbakteriler: Yapısal olarak prokaryot organizmalar olan bu
hücreler, genellikle sert bir hücre duvarı ile çevrelenmiş hücre
zarına sahiptirler. Fakat hücre içi organellerine sahip değildirler. Bir
tane ve genellikle halkasal kromozoma sahiptirler. Tek veya çok
hücreli oloabilirler. E. coli en iyi çalışılmış bir öbakteridir.
Prokaryotlar tipik olarak 1-10 µ çapında ve hayvan
bağırsaklarından asidik sıcak sulara kadar hemen hemen her
koşulda yaşayabilen en basit canlı hücrelerdir.
Bir prokaryotun yüzeyi, diğer hücrelere ve yüzeylere tutunmayi
sağlayan, pili ve hücrenin kendi etrafında dönmesi ve yüzmesini
sağlayan flagella(kamçı) bulundurabilir.
Öbakteriler: Yapısal olarak prokaryot organizmalar olan bu
hücreler, genellikle sert bir hücre duvarı ile çevrelenmiş hücre
zarına sahiptirler. Fakat hücre içi organellerine sahip değildirler. Bir
tane ve genellikle halkasal kromozoma sahiptirler. Tek veya çok
hücreli oloabilirler. E. coli en iyi çalışılmış bir öbakteridir.
Prokaryotlar tipik olarak 1-10 µ çapında ve hayvan
bağırsaklarından asidik sıcak sulara kadar hemen hemen her
koşulda yaşayabilen en basit canlı hücrelerdir.
Bir prokaryotun yüzeyi, diğer hücrelere ve yüzeylere tutunmayi
sağlayan, pili ve hücrenin kendi etrafında dönmesi ve yüzmesini
sağlayan flagella(kamçı) bulundurabilir.
Arkea: Ekstrem çevre koşullarında yaşamaya eğilimlidirler. Yapısal
olarak öbakterilere benzerler. Ancak ribozomal RNA (rRNA)
molekülleri temel alındığında ökaryotlarından ve öbakterileriden
farklılık gösterirler ve zar lipidlerinde ester bağları yerine eter
bağları bulundururlar.
Arkea’lar enerji üretimi ve metabolizmaları yönüyle
karşılaştırıldığında daha çok öbakterilere, replikasyon,
transkripsiyon ve translasyon bakımından ise daha çok ökaryotlara
benzerlik gösterirler.
Ökaryotlar: Mantar, bitki ve hayvan hücreleri lipid zarlarla çevrili
çok tanımlanmış organellere sahiptir.
Özelleşmiş fonksiyonları yerine getirmek üzere yapısal olarak
zarlarla çevrili organellere sahip olmalarıyla tanımlanırlar.
10-100 µ çapındadırlar.
Bitkiler ve birçok mantar aynı zamanda sert bir hücre duvarına
sahiptirler.
Oldukça organize bir jel kıvamındaki sitoplazma, organellere ve
ribozomlara ek olarak, hücrenin hareketini ve şeklini kontrol eden
ve hücrenin bir çok metabolik reaksiyonunun gerçekleşmesine
yardımcı olan hücre iskeleti olarak adlandırılan protein iplikçikleri
demeti bulundurur.
Bu iplikçikler tubilin’den oluşan mikrotubül’leri ve aktinden oluşan
mikroflamentler’i içerir.
Çoğu ökaryotlar organizmanın tüm özelleşmiş dokularını meydana
getirmek için gelişim sırasında farklılaşmaya giren hücrelerden
meydana gelirler.
Farklılaşma: Bir hücre bölündüğünde yavru hücreler her yönden
birbirinin aynısı olabilir veya ana hücreden fonksiyonel olarak
farklılaşmak için gen ifade tarzlarını değiştirebilirler.
Prokaryotlar ve basit yapılı ökaryotlarda spor oluşumu böyle bir
hücresel farklılaşmaya örnektir.
Kompleks çok hücreli ökaryotlarda ise embriyonik hücreler kas,
beyin, karaciğer ve böbrek gibi oldukça özelleşmiş hücrelere
dönüşürler.
DNA içeriği aynı kalmakta fakat anlatıma uğrayan genler
değişmektedir.
A2. ORGANELLER
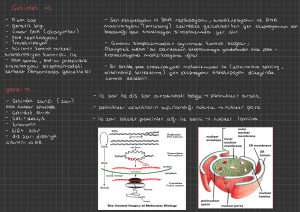
Çekirdekler (Nukleus): Ökaryotik hücre çekirdekleri çoklu
kromozomlar üzerinde hücrenin genetik bilgisini taşır, her bir
kromozom bir tek DNA molekülünden meydana gelir.
Çekirdek nuklear zar olan iki katlı lipit zar ile çevrili olup; bu yapı
oldukça büyük moleküllerin geçebildiği büyük porlar içermektedir.
RNA’nın transkripsiyonu çekirdekte gerçekleşir ve işlenmiş RNA
molekülü translasyonun gerçekleşeceği sitoplazmaya geçer.
Çekirdekçikler (nukleolus) rRNA’nın sentezlendiği ve ribozomların
kısmen toparlandığı çekirdek içerisindeki cisimlerdir.
Mitokondri ve Kloroplastlar: Hücresel solunum, yani besinlerin
adenozin 5’-trifosfat (ATP) formunda enerji meydana getirmek için
oksitlenmesi mitokondrilerde gerçekleşir.
Mitokondriler düz bir dış zara ve kristalar olarak adlandırılan
çıkıntılı iç zara sahiptirler.
Mitokondriler küçük halkasal DNA molekülü, mitokondriye özgü
RNA ve bazı mitokondriyal proteinleri sentezleyen ribozomları
bulundururlar.
Ancak mitokondri (ve kloroplast) proteinlerinin çoğu çekirdekteki
DNA tarafından kodlanır ve sitoplazmada sentezlenir.
Bitkilerin kloroplastları, karbonhidrat ve oksijen meydana
getirmesi için CO2 ve suyun ışık-bağımlı asimilasyonunun
gerçekleştiği fotosentetik merkezlerdir.
Kloroplastlarda kristalar yerine iç zar boşluğunda üçüncü bir zar
sistemine (tilakoidler) sahip olmaları dışında mitokondrilerle
benzer yapılardır.
Endoplazmik retikulum: Sitoplazma içinde yoğun bir zar
sistemidir ve çekirdek zarı ile devamlılığı vardır.
Birçok hücrede 2 form olarak görülürler.
Düz endoplazmik retikulum zarları, bazı lipitlerin biyosentezinden
ve ilaçlar gibi yabancı maddelerin oksidasyon ve detoksifikasyon
(ksenobiyotikler) sorumlu olan birçok enzimi taşır.
Granüllü endoplazmik retikulum (GER) birçok ribozomu
bulundurduğu için bu adı alır. BU ribozomlar, plazma ve süt
proteinleri gibi hücre tarafından salgılanacak özel proteinleri veya
plazma zarı ya da özel organellerin ihtiyacı olan proteinleri
sentezlerler.
GER üzerinde sentezlenen lipitler ve proteinler özelleşmiş taşıma
vesikülleri içerisinde, daha ileri modifikasyon ve ayrıştırmaların
gerçekleştiği ve yassılaşmış zar kesecikleri demeti olan golgi
kompleksine taşınır.
Mikrocisimcikler: Lizozomlar golgi kompleksinin
tomurcuklanarak ayrılan; proteinler, nukleik asitler, lipitler ve
karbonhidratları parçalayacak çeşitli enzimleri içeren küçük, zarla
çevrili organellerdir.
Bunlar hücre dışından alınan veya hasar görmüş organellerden
gelen makromoleküllerin geri dönüşüm merkezleri olarak işlev
görürler.
Oldukça yüksek reaktif serbest radikaller ve hidrojen peroksit
meydana getiren bazı metabolik reaksiyonların ürünleri
peroksizomlar adı verilen organellerde, hücresel bileşenlerin zarar
görmesini önlemek amacıyla etkisizleştirirler.
Peroksizomlar hidrojen peroksit (H2O2) yıkan katalaz enzimini
içerirler.
Glioksizomlar, glikosilat döngüsü reaksiyonlarını gerçekleştiren
özelleşmiş bitkisel peroksizomlardır. Lizozom, peroksizom ve
glioksizomların tümü mikrocisimcikler olarak adlandırılır.
A3. MAKROMOLEKÜLLER
Proteinler ve Nukleik asitler: Proteinler, peptit bağları ile
birbirine bağlanmış amino asitlerin polimerleridir.
DNA ve RNA nukleik asitleri; azotlu baz, 5 karbonlu şeker (Riboz,
deoksiriboz) ve bir fosforik asitten meydana gelen nukleotitlerin
polimeridir.
Hücresel RNA’nın başlıca 3 tipi vardır: mesajcı RNA (mRNA),
ribozomal RNA (rRNA) ve transfer RNA (tRNA).
Nukleik asitler genetik bilginin saklanması ve işlenmesine katılır,
fakat bu bilginin ifadesi için proteinlere ihtiyaç vardır.
Polisakkaritler: Basit şekerlerin glikozidik bağlarla kovalent
olarak bağlanmış polimerleridir.
Selüloz ve nişasta bitkilerde bol bulunan bileşenlerdir.
Bunların her ikiside glukoz polimerleri olmalarına rağmen, glukoz
monomerleri birbirlerine farklı yolla bağlanmışlardır.
Selüloz, α (1-4) bağları ile düz bir polimerdir ve bitki hücre
duvarının başlıca bileşenidir.
Nişasta bir şeker deposudur ve metabolizmada hızlı bir şekilde
hidrolize olarak glukoza dönüşebilen büyük hücre içi tanecikler
olarak bulunur.
Nişasta 2 bileşeni içerir.: α(1-4) bağı doğrusal polimer olan
α-amiloz ve ilave α(1-6) bağlantılarla dallanmış bir polimerdir.
Mantarlar ve bazı hayvansal dokular (karaciğer ve kas) glukozu
amilopektin gibi dallı polimer olan glikojen halinde depo ederler.
Kitin, böceklerin ve kabukluların (Crustacea) dış iskeletinde ve
mantarların hücre duvarlarında bulunur.
Kitin selüloza benzer, fakat monomer birimi
N-asetilglukozamin’dir.
Mukopolisakkaritler, bağ dokusunun fibröz proteinleri içine gömülü
halde jel benzeri solüsyonlar oluştururlar.
Lipitler: Tek başlarına tam anlamıyla makromoleküller olmamakla
birlikte, birçoğu küçük monomerik birimlerden oluşur ve diğer
birçok makromoleküllerin birleştirilmesine katılırlar.
Büyük lipit molekülleri doğada genellikle hidrokarbon formundadır
ve suda çok az çözünürler.
Lipitlerin bir bölümü enerjinin transferi ve depolanmasında görev
yaparken diğerleri ise zarların, koruyucu zarların ve diğer hücre
yapılarının anahtar bileşenidir.
Gliseritler bir mol gliserole esterlenmiş uzun zincirli bir, iki veya üç
yağ asidine sahiptir.
Hayvansal trigiliseritlerde, yağ asitleri çift bağ bulundurmaz
(doymuş) ve bu yüzden zincirler düzdür, moleküller sıkı bir şekilde
paketlenebilir ve sonuçta meydana gelen yağlar katıdır.
Bitkisel yağlardaki yağ asitleri bir veya daha fazla sayıda çift
bağlara sahiptir. Bu zincirlerin açılı yapıları sıkı paketlenmeyi önler,
dolayısıyla bitkisel yağlar oda sıcaklığında sıvı olma eğilimindedir.
Zarlar gliserol ile esterleşmiş iki yağ asiti ile fosforik asitten
meydana gelen fosfolipitleri içerirler.
Ayrıca fosfat genellikle serin, etanolamin, inozitol veya kolin gibi
bir küçük molekülle esterleşirler.
Zarlar ayrıca bir amit bağıyla yağ asidine bağlanmış uzun zincirli
amino alkol sfingozin olan seramit gibi sfingolipitler de bulundurur.
Fosfokolinin bir seramite bağlanması sifingomiyelini üretir.
Kompleks makromoleküller: Birçok makromolekül büyük
biyomoleküllerin birden fazlasının kovalent veya kovalent olmayan
birleşmelerine sahiptir.
Örneğin, hemen hemen tüm enzimler proteindir, fakat bazıları
katalitik aktivite için esansiyel olarak RNA bileşenine kovalent
olmayan bağlarla bağlanması gerekir.
Protein ve nukleik asitten oluşan yapılar nukleoprotein olarak
bilinir.
Ökaryotik kromozom uçlarının replikasyonundan sorumlu olan
telomerazlar ve tRNA’yı olgunlaştıran ribonukleaz P bunlara
örnektir.
Glikoproteinler, protein ve karbonhidrat bileşenlerini içerir.
Proteoglikanlar (mokoproteinler) proteinlerin büyük
kompleksleridir ve mukopolisakkaritler bakteriyel hücre
duvarlarında ve bağ dokusundaki hücrelerarası boşluklarda
bulunur.
Lipoproteinlerde, lipit ve proteinlerin kovalent olmayan bağlarla
bağlanmasıyla oluşmuşlardır.
Çünkü lipitler suda çok az çözünür ve dolayısıyla kanda
lipoproteinler olarak taşınırlar.
A4. MAKROMOLEKÜLER DÜZENLEMELER
Protein kompleksleri: Hücre iskeleti hücrelerin hareketini,
şeklini ve organellerin hücre içi dağılımını organize eden bir
protein filamenti demetidir.
Mikrotubüller, globüler bir protein olan tubulinin uzun
polimerleridir.
Bunlar kamçılama hareketi ile veya sıvıyı hücre yüzeyi boyunca
iterek hücreyi hareket ettiren ve birçok ökaryotik hücrenin
yüzeyinde bulunan tüye benzer siller ve kamçıların, ve hücre
iskeletinin önemli bileşenleridir.
Mikrofilamentler, kasılan yapıları oluşturarak sitoplazmik harekete
neden olan aktin ve miyozin proteinlerinden meydana gelir.
Aktin ve miyozin aynı zamanda kas tellerinin başlıca bileşenleridir.
Nukleoprotein: Ribozomlar, protein sentez yeri olan büyük
sitoplazmik ribonukleoprotein komplekslerdir.
Bakteriyel 70S ribozomları toplam kütlesi 2.5x106 Da olan büyük
(50S) ve küçük (30S) alt ünitelere sahiptir.
50S alt birim 23S ve 5S RNA molekülleri ile 31 farklı protein
içerirken, 30S alt ünite bir 16S RNA molekülü ve 21 farklı proteine
sahiptir.
Ökaryotik 80S ribozomlar 60S (28S, 5.8S ve 5S RNA) ve 40S
(18S RNA) alt ünitelere sahiptir.
Kromatin ökaryotik kromozomları meydana getiren materyaldir.
Kromatin, yaklaşık olarak eşit miktarda DNA ve histon adı verilen
küçük bazik proteinlerden meydana gelmiş bir
deoksiribonukleoprotein kompleksidir.
Bu yapılar nukleozom denilen tekrarlayan birimleri oluşturur.
Nukleozomların doğru bir şekilde toparlanması ve diğer
kompleksleri için toparlayıcı proteinlere ya da kaperonlara
gereksinim vardır.
Histonlar DNA’daki şeker-fosfat omurgasının negatif yükleri
arasındaki itici güçleri nötralize eder ve DNA’nın kromozomlar
içinde sıkı bir şekilde paketlenmesini sağlar.
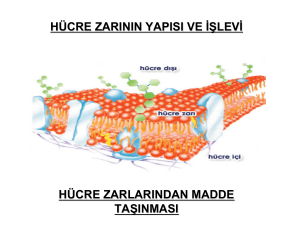
Zarlar: Fosfolipitler ve sfingolipitler sulu ortamda bulunduklarında
doğal olarak dışa bakan kısımında polar grupların, iç kısımda ise
polar olmayan hidrokarbon zincirlerin yer aldığı bir bilayer lipit
katmanı oluşur.
Proteinler hücre diğer önemli bir bileşenidir.
Periferal zar proteinleri dış yüzeye gevşek olarak bağlıdır veya bir
lipit üzerine çökmüş olarak ya da glikozil fosfatidillinozitol çapası
ile asılı bir şekilde bulunurlar.
İntegral zar proteinleri zar içerisine gömülüdür ve zar
parçalanmadan zardan uzaklaştırılamazlar.
Bazıları zarın dış veya iç yüzeyinin dışına çıkarken, transmembran
proteinleri ise bilayer tabaka içerisine tamamen yayılmışlardır ve
hem hücre dışı hem de hücre içi yapılara sahiptirler.
Zar proteinlerinin değişik fonksiyonları vardır.
• Hormonlar ve nörotransmitterler gibi sinyal molekülleri için
reseptörler
• Hücre dışı moleküllerin hücre içine alımından önce yıkımı için
enzimler
• Küçük, polar iyonlar ve moleküülerin seçici taşınımı için polar
veya kanallar
• Hücre-hücre etkileşimi için aracılar
B. PROTEİN YAPISI
B1. AMİNO ASİTLER
Yapısı: Proteinler L-amino asitlerin polimerleridir. Prolin hariç
tutulursa, proteinlerde bulunan tüm 20 aa’in hepsi genel olarak
α-karbonuna bağlı bir karboksil grup, bir primer amino grubu, bir
proton ve her bir aa’e farklı olan bir yan zincir ( R ) sahiptir.
Glisin dışında α-karbon atomu asimetriktir. Bu sayede aa’ler
optikçe aktif stereoizomer çiftleri şeklinde var olabilir (D- ve L-).
Buna rağmen, proteinlerde sadece L-izomerleri bulunur.
Aa’ler sulu çözeltilerde dipolar iyonlardır (zwitteriyonlar) ve hem
asit hemde baz gibi davranırlar (amfoterik).
Yüklü yan zincirler: Referans noktası olarak pH 7 alınırsa, bazı
aa’lerin yan zincirleri bu ph’da ekstra bir pozitif veya negatif yük
sağladığından dolayı iyonlaşabilen gruplara sahiptir.
Asidik aa’ler, aspartik asit ve glutamik asit, genellikle iyonlaşan
(negatif yüklü) ilave karboksil gruplarına sahiptir.
Bazik aa’ler, pozitif yüklü gruplara sahiptir; lizin ε-karbon atomuna
bağlı ikinci bir amino grubuna sahipken, arjinin bir guanidin
grubuna sahiptir.
Polar yüksüz yan zincirler: Bunlar suyla hidrojen bağlarını
oluşturan grupları içerirler.
Bunlar yüklü aa’lerle birlikte genelde hidrofilik (su seven) olarak
tanımlanırlar.
Serin ve treonin hidroksil gruplarına sahipken, asparagin ve
glutamin, aspartik asit ve glutamik asitin amit türevleridir.
Polar olmayan alifatik yan zincirler: Glisin yan zincir yerinde
bir hidrojen atumuna sahiptir ve optikçe inaktiftir.
Prolin sekonder aa olmasından dolayı genel aa’lerden ayrı özellik
gösterir.
Alanin, valin, lösin ve izolösin yan zincirlerinde hidrofobik (su
sevmeyen) alkil gruplarına sahiptir ve bu gruplar protein
yapılarında hidrofobik etkileşimlere katılırlar.
Aromatik yan zincirler: Fenilalanin, tirozin ve triptofan çok
büyük hidrofobik yan zincirlere sahiptir.
Bu aa’lerin aromatik yapıları, 280 nm’de maksimum absorbsiyon
yapan proteinlerin, UV absorbansının büyük bir kısmını
gerçekleştirir.
B2. PROTEİN YAPISI VE FONKSİYONU
Boyutları ve şekilleri: Proteinler iki büyük gruba ayrılabilirler.
Globüler proteinler sıkı bir şekilde katlanmıştır ve solüsyonlarda az
veya çok küresel partiküller halindedirler. Enzimlerin çoğu doğada
globüler yapıdadır.
Fibröz proteinler oldukça büyük aksiyal oranlara (uzunluk/genişlik)
sahiptir ve bunlar önemli yapısal proteinlerdir, saç keratini gibi.
Primer yapı: Bir aa’in α-karboksil grubu diğer bir aa’in α-amino
grubuna yaygın olarak peptit bağı olarak bilinen bir amit bağı ile
kovalent olarak bağlanır.
İki aa’in bu yolla birleşmesi ile oluşan ürün bir dipeptittir. Çok
sayıda aa peptit bağları ile bağlanarak bir polipeptidi oluşturur.
α-karbon atomlarının tekrarlayan dizisi ve peptit bağları
polipeptidin omurgasını oluştururken, aa’lerin farklı yan zincirleri
proteinlerde fonksiyonelliği belirler.
Aa’lerin N-ucundan C-ucuna olan dizisi polipeptidin primer
yapısıdır.
Sekonder yapı: Peptit bağlarının birleşme yerlerinde serbest
rotasyon olmasına rağmen bu durum, peptit bağının düz ve
serbest yapısını ortaya çıkarır.
Bu polarite uygun aralıkta ve yöndeki peptit bağ üniteleri arasında
H bağlarının oluşmasına neden olur.
Bu sayede, H bağları ile bir arada tutulan birkaç düzenli yapılar
şeklinde polipeptit zincirleri katlanabilir.
En iyi bilinen sekonder yapı α-heliks’dir.
α-heliks sekonder yapı örnekleri genelde globüler proteinlerde, az
bir kısmı fibröz proteinlerde bulunur.
Tersiyer yapı: α-heliks ve diğer az rastlanan sekonder yapılar üç
boyutlu düzenlenerek polipeptidin tersiyer yapısını oluşturur.
Uygun şartlarda çoğu polipeptidler genelde çok az bir enerji ile
doğru tersiyer yapıyı oluşturacak şekilde katlanırlar.
Fakat, yeni polipeptit zincir sentezleri öncesi yanlış katlanmaları
önlemede yardımcı olan kaperonlar adı verilen proteinlerin
katkısıyla in vivo doğru katlanmalar yapılır.
Yüksek sıcaklık şoku veya aşırı pH uygulaması ile sekonder ve
tersiyer yapıların bozulması, proteinlerin denatürasyonuna neden
olur ve rastgele sarmal yapısını ortaya çıkarır.
Kuaterner yapı: Proteinlerin çoğu iki veya daha fazla polipeptit
zincirinden meydana gelmiştir.
Bu zincirler birbirinin aynısı veya farklısı olabilir.
Hemoglobin iki α globin ve iki β-globin zincirine (α2β2) sahiptir.
Tersiyer yapıyı kararlı hale getiren benzer kuvvetler, bu alt
üniteleri bir arada tutar.
Bunlara polipeptitdeki sisteinler arasındaki disülfit bağları da
dahildir.
Bu organizasyonun seviyesi kuaterner yapı olarak adlandırılır ve
belirli sonuçlara sahiptir.
Proteinler ne yapar?
• Enzimler; Katalitik olarak aktif birkaç RNA molekülü hariç
bütün enzimler proteindir.
Biyokimyasal reaksiyonların hızını önemli ölçüde arttırırlar.
Substrata bağlanma, van der Waals kuvvetleri, H bağları, tuz
köprüleri ve hidrofobik kuvvetleri kapsayan spesifik Aa yan
zincirleri ile kovalent olmayan etkileşimlerle gerçekleşir.
* Sinyalleşme; Hücre zarındaki reseptör proteinler ligandlara
(örn. hormonlar) bağlanabilir.
Bu durumda yapısal değişiklikler ortaya çıkar, hücre içerisinde
liganda cevap olarak reaksiyonlar başlar.
• Taşıma ve depolama; Hemoglobin kırmızı hücrelerde oksijen
taşırken, transferrin ise karaciğere demir taşır.
Demir karaciğerde ferritin proteinlerine bağlı olarak depolanır.
Besinsel yağlar kanda lipoproteinler tarafından taşınır.
• Yapı ve hareket; Deri, kemik ve bağ dokudaki başlıca protein
kollojendir. Saçlar ise esas olarak keratinden meydana
gelmiştir.
Hücre iskeleti örneğinde olduğu gibi hücre içerisinde daha birçok
yapısal proteinler bulunmaktadır. Başlıca kas proteinleri olan aktin
ve miyozin kas kasılmasını sağlayan filamentleri oluştururlar
• Beslanme; Kazein ve ovalbumin süt ve yumurtanın başlıca
proteinleridir ve gelişmekte olan yavrunun büyümesi için
gerekli Aa’leri sağlamada kullanılırlar.
• Bağışıklık; Bakteri, virüs diğer yabancı materyalleri (antijen)
tanıyan ve onlara bağlanan antikorlar proteindir.
* Regülasyon; Transkripsiyon faktörleri DNA’ya bağlanırlar ve
DNA’nın fonksiyonlarını ayarlarlar.