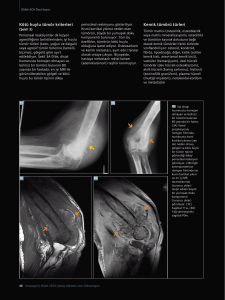
özge ateşli.tez - Erciyes Üniversitesi Eczacılık Fakültesi
advertisement

1 T.C. ERCİYES ÜNİVERSİTESİ ECZACILIK FAKÜLTESİ MONOKLONAL ANTİKORLARLA KANSER TEDAVİSİ Hazırlayan Özge ATEŞLİ Danışman Yrd. Doç. Dr. Dilşad ONBAŞILI Farmasötik Biyoteknoloji Anabilim Dalı Bitirme Tezi Mayıs 2013 KAYSERİ i BİLİMSEL ETİĞE UYGUNLUK Bu çalışmadaki tüm bilgilerin, akademik ve etik kurallara uygun bir şekilde elde edildiğini beyan ederim. Aynı zamanda bu kurallar ve davranışların gerektirdiği gibi, bu çalışmanın özünde olmayan tüm materyal ve sonuçları tam olarak aktardığımı ve referans gösterdiğimi belirtirim. Özge ATEŞLİ ii YÖNERGEYE UYGUNLUK “Monoklonal antikorlarla kanser tedavisi”adlı bitirme ödevi Erciyes Üniversitesi Lisansüstü Tez Önerisi ve Tez Yazma Yönergesi’ ne uygun olarak hazırlanmıştır. Hazırlayan Özge ATEŞLİ Danışman Yrd. Doç. Dr. Dilşad ONBAŞILI Farmasötik Biyoteknoloji Anabilim Dalı Başkanı Yrd. Doç. Dr. Dilşad ONBAŞILI iii “Monoklonal antikorlarla kanser tedavisi”adlı Bitirme Ödevi Erciyes Üniversitesi Lisansüstü Tez Önerisi ve Tez Yazma Yönergesi’ne uygun olarak hazırlanmış ve Farmasötik Biyoteknoloji Anabilim Dalında Bitirme Ödevi olarak kabul edilmiştir. Tezi Hazırlayan Özge ATEŞLİ Danışman Yrd. Doç. Dr. Dilşad ONBAŞILI Farmasötik Biyoteknoloji Anabilim Dalı Başkanı Yrd. Doç. Dr. Dilşad ONBAŞILI ONAY: Bu bitirme ödevinin kabulü Eczacılık Fakültesi Dekanlığı’ nın................... tarih ve …………..……………sayılı kararı ile onaylanmıştır. …/…/…… Prof. Dr. Müberra KOŞAR Dekan iv TEŞEKKÜR Bitirme ödevimi hazırlarken çalışmalarımı yönlendirmesinde, araştırmalarımın her aşamasında bilgi, öneri ve yardımlarını esirgemeyerek akademik ortamda olduğu kadar insani ilişkilerinde de sonsuz desteğiyle gelişmeme katkıda bulunan danışman hocam Sayın Yrd. Doç. Dr. Dilşad ONBAŞILI’ya ve Sayın Arş. Gör. Berrak ALTINSOY’a, çalışmam süresince emek ve desteği için arkadaşım Erhan TURAN’a, yaşamımın her döneminde bana duydukları güven için aileme en derin duygularla teşekkür ederim. Özge ATEŞLİ Kayseri, Mayıs 2013 v MONOKLONAL ANTİKORLARLA KANSER TEDAVİSİ Özge ATEŞLİ Erciyes Üniversitesi, Eczacılık Fakültesi Farmasötik Biyoteknoloji Anabilim Dalı Bitirme Ödevi, Mayıs 2013 Danışman: Yrd. Doç. Dr. Dilşad ONBAŞILI ÖZET Vücut, son derece farklı hücrelerden ve moleküllerden oluşan bir savunma sistemi tarafından korunmaktadır. Bu sistemde yer alan elemanlar öncelikle organizmanın kalıtsal yapısına yabancı, antijen olarak isimlendirilen her türlü hücre dışı madde ve mikroorganizmanın vücuda girmesini engellemektedir. Antikorlar ise antijenik uyarım sonucu B hücrelerinin değişimi ile oluşan plazma hücreleri tarafından sentezlenen antijenleri bularak onların yok edilmesini sağlayan immünoglobulinler olarak adlandırılmaktadırlar. Hibridoma teknolojisinin gelişmesiyle monoklonal antikorlar gerek tanı gerekse tedavi konusunda özellikle de kanserde yeri doldurulamaz hale gelmiştir. Monoklonal antikorlar tümör hücrelerindeki spesifik antijenlere karşı üretilmektedirler ve bu özellikleriyle hedeflenmiş silahlara benzetilmektedirler. Monoklonal antikorlar eczacılıkta hastalıkların teşhisi, tedavisi ve profilaksisi için kullanılmaktadır. Son 20 yıl içinde onlarca monoklonal antikor, ya tümör-spesifik immün cevapları geliştiren veya aktive eden ya tümör hücre içi sinyal iletim aşamalarını bozan ya özellikle malign hücrelere toksin dağıtan ya da tümör-stroma etkileşimini bloke eden bu dolaylı antineoplastik etki kapasiteleri için geliştirilmiş ve karakterize edilmiştir. Bu derlemede amaç, monoklonal antikorların kanser tedavisindeki kullanımının araştırılmasıdır. Anahtar Kelimeler: Monoklonal antikor, kanser tedavisi, alemtuzumab, rituksimab, trastuzumab vi MONOCLONAL ANTIBODIES IN CANCER THERAPY Özge ATEŞLİ Erciyes University, Faculty of Pharmacy Department of Pharmaceutical Biotechnology Graduation Project, May 2013 Advisor: Yrd. Doç. Dr. Dilşad ONBAŞILI ABSTRACT Body is protected by a defense system which is formed ultra different cells and molecules. Elements which are hold a place in this system primarily prevent from entering all kinds of extracellular subtance and microorganism known as antigen which is foreign to the structure of hereditary organism into the body. Antibodies which find antigens and provide destruction of them are called as immunoglobulins which is synthesized by plasma cells which are formed with change of B cells as a result of antigenic stimulation. With the development of hybridoma technology, monoclonal antibodies have become an irreplaceable especially in the diagnosis and treatment of cancer. Monoclonal antibodies are produced against tumor-specific antigens and are liken to targeted weapons. Monoclonal antibodies are used for diagnosis, treatment and prophylaxis of diseases at pharmacy. During the past 20 years, monoclonal antibodies have been developed and characterized for their capacity to mediate antineoplastic effects, either as they activate/enhance tumor-specific immune responses, either as they interrupt cancer cell-intrinsic signal transduction cascades, either as they specifically delivery toxins to malignant cells or as they block the tumor-stroma interaction. In this review the purpose is researching usage of monoclonal antibodies in cancer therapy. Key Words: Monoclonal antibody, cancer therapy, alemtuzumab, rituximab, trastuzumab vii İÇİNDEKİLER BİLİMSEL ETİĞE UYGUNLUK .................................................................................. i YÖNERGEYE UYGUNLUK......................................................................................... ii KABUL ONAY ...............................................................................................................iii TEŞEKKÜR ................................................................................................................... iv ÖZET................................................................................................................................ v ABSTRACT .................................................................................................................... vi İÇİNDEKİLER ............................................................................................................. vii TABLOLAR LİSTESİ ................................................................................................... ix ŞEKİLLER LİSTESİ ...................................................................................................... x KISALTMALAR ........................................................................................................... xi 1. GİRİŞ VE AMAÇ ....................................................................................................... 1 2. GENEL BİLGİLER .................................................................................................... 4 2.1.LENFOİD SİSTEM ve İMMÜN CEVAP ............................................................... 4 2.1.1.Lenfositler ......................................................................................................... 5 2.1.1.1.B Lenfositler............................................................................................... 5 2.1.1.2.T Lenfositler ............................................................................................... 5 2.1.1.2.1.Sitotoksik T lenfositleri ....................................................................... 6 2.1.1.2.2.Yardımcı T lenfositler ......................................................................... 6 2.1.1.3.Doğal Öldürücü Hücreler (Naturak killer=NK) ........................................ 7 2.1.2.Mononükleer fagositik sistem........................................................................... 7 2.1.3.Polimorfonüklear granülositler ......................................................................... 9 2.1.3.1.Nötrofiller................................................................................................... 9 2.1.3.2.Bazofiller ve Mast Hücreleri ...................................................................... 9 2.1.3.3.Eozinofiller................................................................................................. 9 2.1.4.Major Histokompatibilite Kompleksi ............................................................. 10 2.1.4.1.Sınıf-I Molekülleri ................................................................................... 10 viii 2.1.4.2.Sınıf-II Molekülleri .................................................................................. 10 2.1.4.3.Sınıf-III Molekülleri ................................................................................. 11 2.1.5.Kompleman Sistemi (Tamamlayıcı Sistem) ................................................... 11 2.1.6.Sitokinler......................................................................................................... 13 2.2. TÜMÖR İMMÜNOLOJİSİ .................................................................................. 17 2.2.1. Tümör Antijenleri .......................................................................................... 18 2.2.2. Efektör Mekanizmalar ................................................................................... 18 2.2.3. Kanserin İmmün Sistemden Kaçış Mekanizmaları ....................................... 20 2.3.ANTİKORLAR ..................................................................................................... 20 2.3.1. Monoklonal Antikor ...................................................................................... 22 2.3.2. Monoklonal Antikorlar ile Kanser Tedavisi .................................................. 28 3. TARTIŞMA VE SONUÇ.......................................................................................... 36 KAYNAKLAR .............................................................................................................. 41 ÖZGEÇMİŞ ................................................................................................................... 46 ix TABLOLAR LİSTESİ Tablo 2.1. mAb veya mAb ile ilişkili terapötikler için uluslararası son ekler ............... 27 Tablo 2.2. Günümüzde kanser tedavisi için onaylı monoklonal antikorlar.................... 33 Tablo 2.3. Faz III-IV çalışmalarda kanser tedavisi için değerlendirme aşamasında olan monoklonal antikorlar .................................................................................. 34 x ŞEKİLLER LİSTESİ Şekil 2.1. Bağışık yanıtta rol alan hücrelerin etkileşimi .................................................. 7 Şekil 2.2. Klasik ve alternatif kompleman yollarının aktivasyonları ve membran atak kompleksi (C5b-9) oluşumunun basamakları .............................................. 12 Şekil 2.3. GM-CSF ve T hücre yanıtları ........................................................................ 16 Şekil 2.4. İmmün sistemin doğuştan ve kazanılmış bölümlerinin ana bileşenlerinin ve hedeflerinin şematik özeti ............................................................................... 17 Şekil 2.5. Antikorun yapısı ............................................................................................ 21 Şekil 2.6. Monoklonal antikorların üretimi. ................................................................... 26 Şekil 2.7. Panitumumab’ın etki mekanizması................................................................. 28 Şekil 2.8. Tümör hücreleri, stroma hücreleri, perisitler ve endotel hücreleri arasındaki intraselüler sinyalizasyon ............................................................................. 29 Şekil 2.9. CD20 monoklonal antikorlarının etki mekanizmaları .................................... 30 Şekil 2.10. Katumaksomab’ın öne sürülen etki mekanizması ........................................ 31 xi KISALTMALAR ADCC : Antikor bağımlı hücresel sitotoksisite ADCP : Antikor bağımlı hücresel fagositoz AIDS : Edinilmiş yetersiz bağışıklık sistemi sendromu CDC : Kompleman bağımlı sitotoksisite CMV : Sitomegalovirüs CTL : Sitotoksik T lenfositleri DC-CK : Dendritik hücre sitokini DNA : Deoksiribonükleik asit EGF : Epitelyal büyüme faktörü EGFR : Epitelyal büyüme faktörü reseptörü EpCAM : Epitelyal hücre adhezyon molekülü Fab : Antikorun antijen bağlama parçası Fc : Antikorun sabit parçası FcγR : Fc γ reseptörü FcRn : Fc neonatal reseptör GM-CSF : Granülosit-makrofaj koloni uyarıcı faktör HAT : Hipoksantin, Aminopterin, Timidin HER : İnsan büyüme faktörü reseptörü HGPRT : Hipoksantin-guanin fosforiboziltransferaz HLA : İnsan lökosit antijenleri IFN : İnterferon xii Ig : İmmünoglobulin IGFR : İnsülin benzeri büyüme faktörü IL : İnterlökin ITAM : İmmünoreseptör tirozin bazlı aktivasyon motifleri ITIM : İmmünoreseptör tirozin bazlı inhibitör motifleri LAK : Lenfokinle aktive edilmiş öldürücü hücreler LGL : İri granüllü lenfositler mAb : Monoklonal antikor MAC : Membran atak kompleksi M-CSF : Makrofaj koloni uyarıcı faktör MHC : Major histokompatibilite kompleksi NK : Doğal öldürücü hücreler NO : Nitrik oksit NSCLC : Küçük hücreli olmayan akciğer kanseri O2 : PDGF : Platelet kaynaklı büyüme faktörü PEG : Polietilen glikol PET/CT : Pozitron emisyon tomografisi/bilgisayarlı tomografi PMNL : Polimorfonüklear lökosit RANKL : Reseptör aktifleştirici nükleus kappa ligandı RNA : Ribonükleik asit RIT-Z : Zevalin ile radyoimmünoterapi Oksijen xiii SCLC : Küçük hücreli akciğer kanseri TCGF : T hücresi büyüme faktörü TCR : T hücre reseptörü TGF : Değişiklik yapan büyüme faktörü TNF : Tümör nekrozis faktör VEGF : Vasküler endotelyal büyüme faktörü 1 1. GİRİŞ VE AMAÇ Doğadaki bütün canlılar kendilerinden olmayan doku, hücre ve moleküllere karşı savunma sistemlerine sahiptirler. Örnek olarak bakteriler fajlara (bakteri virüslerine) karşı kendilerini bu saldırganların DNA ve RNA’larını parçalayan enzimler üreterek korumaktadırlar. Yüksek canlılarda ise çok daha karmaşık bir bağışıklık sistemi bulunmaktadır. Omurgalılarda bağışıklık sistemi çok sayıda farklı hücre ve molekül içermektedir (1). Bağışıklık eski çağlardan bu yana insanların ilgi noktası olmuş ve bazı bağışıklama yöntemleri geliştirilmiştir. Örneğin Türkler, çiçek hastalığı geçirenlerin yaralarındaki kabukları toz haline getirerek burunlarından çekip ya da kollarını ateşe tutulmuş bir bıçakla çizerek oluşturdukları yaraya bu kabukları bulaştırdıkları bilinmektedir. Erlich ve Elie Metchnikoff’un kendilerine 1905 Nobel Ödülü’nü getiren çalışmalarının bağışıklık sisteminin anlaşılmasında öncü niteliğe sahiptirler. Günümüzde bağışıklık sisteminin geniş ölçüde aydınlatıldığı söylenebilir ve bu sistemi oluşturan unsurlardan hastalıkların tanısında, tedavisinde geniş ölçüde yararlanılmaktadır (1). Bu sistemde yer alan elemanlar öncelikle organizmanın kalıtsal yapısına yabancı, antijen olarak isimlendirilen her türlü hücre dışı madde ve mikroorganizmanın vücuda girmesini engellemektedir. Antijenler, organizmaya girdiğinde kendisine karşı bir bağışık yanıt oluşmasına yol açan ve bu yanıt sonucunda oluşan ürün ile özgül olarak birleşebilen maddeler olarak tanımlanmaktadır (1,2). Antijenler, deri, solunum ve sindirim sistemi gibi engelleri aşarak, organizmaya dahil olduklarında, savunma sistemi hemen harekete geçer. Kemik iliği, timus, lenf bezleri ve dalak gibi özelleşmiş merkezlerde yer alan fagositler, makrofajlar, lenfositler (B ve T hücreleri) gibi savunma hücreleri ve molekülleri devreye girerler. İlk aşamada, öncü hücreler olan fagositler ve makrofajlar antijenleri yok etmeye çalışırlar; etkili olamadıkları durumda B hücresi ve T hücresi olarak adlandırılan lenfositler devreye girerler. Antijen varlığını haber alan T 2 hücreleri, savunma sisteminin yönetimini ellerine alarak, diğer hücreleri uyarırlar. Sitotoksik T hücreleri antijenleri yok etmek için uğraş verirken, savunmadan sorumlu B hücreleri de bağışıklığın akıllı molekülleri olarak adlandırılan antikorları sentezlemeye başlarlar. Antikorlar, antijenik uyarım sonucu B hücrelerin değişimi ile oluşan plazma hücreleri tarafından sentezlenen antijenleri bularak onların yok edilmesini sağlayan immünoglobulinler olarak adlandırılmaktadırlar. Bu moleküllerin üstünlüğü, üç boyutlu bir yapıda, tıpkı anahtar-kilit arasındaki uyumu andıracak şekilde, antijene özgün olarak bağlanmaktır. Böylece antikorlar, organizmaya giren antijenlere bağlanarak onları etkisiz hale getirirler veya kompleman sistemini ve diğer savunma hücrelerini harekete geçirerek antijenleri yok ederler (1,3). Hastalıklarla savaşta antikorların büyük miktarlarda ve saf olarak elde edilmesi klasik yöntemlerle oldukça zordur. 1975 yılında Köhler ve Milstein’ın geliştirdiği hibridoma yöntemiyle, hedef yapıda yer alan bir antijenik bölgeye (epitop) karşı limitsiz miktarda monoklonal antikor (mAb) üretmek mümkün olmuştur. Monoklonal antikorlar hibrit hücreler tarafından üretilen homojen antikor topluluklarıdır. Bu yöntemle araştırmacılar, 1984 yılında tıp alanında Nobel ödülünü kazanmışlardır. Bu teknoloji ile, antikorların saf halde ve oldukça büyük miktarlarda üretilmesi olanaklı hale gelmiştir. Bu yöntem basitçe şöyledir: Öncelikle, istenen antikorları doğal olarak üreten hücreler elde edilir. Daha sonra bu hücrelere sonsuz bölünme yeteneği kazandırılır ve kültür ortamında, istenen antikoru üretecek hibrit hücreler geliştirilir. Örneğin, myeloma, kemik iliğinde oluşan ve hücre kültüründe üretilmeye uygun olan bir tümör tipidir. Myeloma hücreleri, antikor üretme yeteneğine sahip olan dalak hücreleri ile kaynaştırıldıklarında, oluşan hücreler büyük miktarlarda mAb üretirler (1,4). Monoklonal antikorlar günümüzde çok çeşitli tanı ve tedavi yöntemlerinde kullanılmaktadırlar. Kanser tedavisi için ilk onayı 1997 yılında alan rituksimab (MabThera®, Rituxan®) tedavide klinik süreci başlatmış ve günümüzde kanser tedavisinde kullanılan mAb’ların sayısı alemtuzumab (MabCampath, Campath-1H®), bevasizumab (Avastin®), brentuksimab vedotin (Adcentis®), katumaksomab (Removab®), setuksimab (Erbitux®), denosumab (Prolia®), gemtuzumab ozogamisin (Mylotarg®), (Arzerra®), ibritumomab panitumumab (Zevalin®), ipilimumab (Yervoy®), ofatumumab (Vectibix®), rituksimab (MabThera®, Rituxan®), tositumomab (Bexxar®) ve trastuzumab (Herceptin®) ile 14’e ulaşmıştır. Bu mAb’lar 3 kronik lenfositik lösemi, glioblastoma multiforme, kolorektal, böbrek ve akciğer kanseri, Hodgkin lenfoma, non-Hodgkin lenfoma, anaplastik büyük hücreli lenfoma, EpCAM pozitif malign assitli durumlar, baş-boyun kanseri, prostat kanseri, melanoma, mide veya gastroözofageal adenokarsinoması gibi kanser tedavilerinde kullanılmaktadırlar. Bu derlemede immün sistem hücreleri de anlatılarak günümüzdeki monoklonal antikorlar ve bu antikorlarla kanser tedavisinin araştırılması amaçlanmıştır. 4 2. GENEL BİLGİLER 2.1.LENFOİD SİSTEM ve İMMÜN CEVAP İmmünite; vücuda giren yabancı maddelerin nötralize edilmesi, dışarıya atılması veya metabolize edilmesi için vücudun geliştirdiği savunma mekanizmasıdır. Mikroorganizmalara, malignensilere ve otoimmün fenomenlere karşı konağın savunma mekanizmalarını içeren bir kompleks sistem olan immün sistem başlıca hücresel immünite, hümoral immünite, fagositik sistem ve kompleman sistemi olmak üzere dört ana unsurdan meydana gelmiştir. İmmün sistemi oluşturan hücrelerin iki ana farklılaşma şekli vardır: • Lenfoid seri: Lenfositleri yapar. • Myeloid seri: Fagositleri ve diğer hücreleri yapar. Lenfoid seride hücresel immüniteden sorumlu T-lenfositler ve hümoral immüniteden sorumlu B-lenfositler olmak üzere fonksiyonları farklı iki ana hücre grubu vardır. Fagositler grubunda ise monositler ve polimorf nükleuslu lökositler (nötrofiller, eozinofiller ve bazofiller) iki temel hücreyi oluştururlar.Bütün bu hücre sistemleri birbirlerine son derece bağlı olmalarına rağmen fonksiyonları farklıdır. Bu iki hücre sistemine ek olarak ‘aksesuar hücreler’ immün cevabın oluşmasında önemli rol oynarlar.Bir antijene karşı immün cevap oluşabilmesi T ve B lenfositleri, monositler ve MHC (major histocompatibility complex) ürünleri arasındaki kompleks bir reaksiyon serisinin gerçekleşmesine bağlıdır. Bunlardan başka diğer hücre tipleri de immün cevabın oluşumuna katılabilmektedir. Bu hücreler arasında dentritik hücreler, NK (natural killer) hücreleri, Langerhans (dendritik) hücreleri ve endotelyal hücreler bulunmaktadır. Lenfoid sistemin lenfoid dokuları primer (santral) ve sekonder (periferik) olmak üzere ikiye ayrılırlar. Santral lenfoid organları kemik iliği, timustur. Periferik lenfoid organlar ise lenf nodları, dalak, tonsiller ve Peyer plakları gibi intestinal lenfoid dokulardır (5). 5 2.1.1.Lenfositler İmmün yanıtta birincil akyuvarlar lenfositlerdir. Kemik iliğindeki kök hücrelerden gelmektedir. Antijen tanıyan ve dolaşımda bulunan uzun ömürlü hücrelerdir. İki ana lenfosit çeşidi B hücreler ve T hücrelerdir (6). Yarı ömrü 5-7 gün gibi kısa ama genellikle daha büyük olanlar B-lenfositler olarak tanımlanırken; küçük ama yarı ömrü aylar ve yıllar gibi uzun olanlar ise T-lenfositler olarak tanımlanırlar (5). Üçüncü tip lenfosit doğal katil hücre (NK-natural killer) dir. Bu tip lenfosit doğuştan gelen bağışıklığın bir parçasıdır. NK hücreleri (bağışıklık tepkisinin ilk evrelerinde) yabancı olarak gördükleri hücreleri karakteristik antijenleri tanımaya gerek görmeden yok eder. Birçok bağışıklık yetersizliği hastalıklarında (AIDS dahil olmak üzere) NK hücreleri anormal çalışır (6). 2.1.1.1.B Lenfositler İki ana B lenfosit çeşidi bulunmaktadır. • İlk çeşit, antijen tarafından uyarılınca plazma hücresine dönüşür. (Plazma hücreleri antijenlerle savaşacak karakteristik antikorları üretir.) • İkinci çeşit, B hafıza (B memory) hücresidir. Bu tip hafıza hücreleri daha önce karşılaşılan antijen bilgisini saklar. Antijen varken, bu hücre kendini klonlar veya bölünerek çoğalır. Bütün yeni oluşmuş hücreler eskiden karşılaşılan antijenler hakkında bilgiye sahiptir. Plazma hücreleri antikor adı verilen proteinleri üretir. Antikorlar, aynı zamanda kan serumundaki immünoglobulin olarak adlandırılırlar (6). 2.1.1.2.T Lenfositler T lenfositler hücresel tipte bağışık yanıttan sorumludur. Kemik iliğinde yapılan T öncü hücreler timusta olgun T lenfosit haline gelirler. Bu olgunlaşma sırasında T lenfosit yüzeyinde pek çok reseptör yerleşir. 1980 sonrası hücre yüzey molekülleri üzerindeki çalışmalar artmış ve pek çok reseptör çeşidi gösterilmiştir. Ancak bunların 6 isimlendirilmesi sorun yaratmış ve karışıklığı önlemek için kan hücreleri ile ilgili yüzey reseptörler numaralanmış ve CD ile ifade edilmişlerdir. CD2, CD4, CD8 gibi (7). T-hücre yüzeyinde yüzey immünglobulin bulunmaz. Bunun yerine antijenleri özgül olarak tanıyan ‘T hücre reseptörü = TCR’ bulunur. Bir T lenfositi sadece tek bir çeşit antijen için TCR taşır ve B lenfositlerinde olduğu gibi immün sistemde zaman içinde karşılaşma ihtimali olan onbinlerce çeşit antijene yanıt verebilecek onbinlerce çeşit T lenfositi bulunur (7). Bağışık yanıttaki rolleri açısından T-hücre topluluğunun homojen (tek tip) olmadığı ve yapı ve işlev özelliği farklı olan alt grupların bulunduğu bilinmektedir. Tüm T lenfositlerde bulunan ortak yüzey molekülleri (CD2, CD3, CD5 gibi) yanında bu alt gruplardaki farklı yüzey moleküller onların ayırdedilmesinde kullanılır (7). T-lenfositler başlıca iki alt gruba ayrılırlar: 2.1.1.2.1.Sitotoksik T lenfositleri (CTL) Bu hücrelerin yüzeyinde CD8 molekülleri bulunur. Organizmaya giren yabancı hücreleri, organizmada şekillenen tümör hücrelerini, virüsle enfekte hücreleri ve vücuda ait bazı hücreleri öldürme yeteneğine sahip bir direkt saldırı hücresidir (8). Sitotoksik T lenfositler, normal olmayan hücreleri kolayca tanıyarak onları ekstraselüler bir mekanizma ile öldürür. Bu hücrelerin fagositoz aktiviteleri yoktur. Bu nedenle intraselüler öldürme yeteneğine sahip değildirler (8). Sitotoksik T lenfositler, antijen sunan hücrelerin ve hedef hücrelerin yüzeylerindeki MHC-I molekülü ile birleşmiş olan endojenik peptid antijenleri kendilerinde bulunan αβ TCR’ler yardımıyla tanıyarak uyarılırlar. Bu uyarımdan sonra sitotoksik T lenfositleri perforin olarak isimlendirilen ve hedef hücrenin membranında nokta şeklinde geniş yuvarlak boşluklar oluşturan maddeler sentezlerler (8). 2.1.1.2.2.Yardımcı T lenfositler T lenfositlerin büyük bir çoğunluğunu yardımcı T lenfositler oluşturmaktadır. Yüzeylerinde CD3 ve CD4 reseptörlerini taşıyan yardımcı T lenfositler, antijen sunan hücreler tarafından işlenerek MHC-II molekülleri ile birlikte hücre yüzeyinde 7 kendilerine sunulan yabancı peptid yapıdaki antijenleri kendilerinde bulunan αβ TCR’ler yardımıyla tanır ve uyarılırlar. Bu uyarımlar sonucunda bir farklılaşma ve gelişme aşamasına geçerek, çeşitli gen düzenlemelerine bağlı olarak sitokin sentezleyecek kapasiteye ulaşırlar (8). Şekil 2.1. Bağışık yanıtta rol alan hücrelerin etkileşimi (7) 2.1.1.3.Doğal Öldürücü Hücreler (Naturak killer=NK) Sitoplazmalarındaki azurofilik granüllerden dolayı ve diğer lenfositlerden daha büyük olmaları nedeniyle iri granüllü lenfositler (LGL=Large granular lymphocyte) olarak da isimlendirilirler. NK hücrelerde bulunan temel CD molekülleri CD16 ve CD56’dır. LGL morfolojisi gösteren dinlenme halindeki NK hücreleri, hedef hücreye bağlandıkları zaman sitolitik granülleri salgılarlar. Hedef hücre membranındaki monomerleri polimerize ederek silindirik transmembran porları oluştururlar ve bu da hedef hücrenin ozmotik lizisine neden olur. Ayrıca NK hücreleri, NK sitotoksik faktör olarak isimlendirilen IL-3, GM-CSF (granülosit-makrofaj koloni uyarıcı faktör), M-CSF, IFNα, β, γ, TNF-α, β gibi eriyebilen sitokinleri de salarlar (9). 2.1.2.Mononükleer fagositik sistem Mononükleer fagositik sistem immün sistemin ikinci büyük hücre grubunu oluşturur ve primer fonksiyonu fagositoz olan hücrelerden oluşur. Mononükleer fagositik sistemin bütün hücreleri kemik iliğinden gelişir. Önce monositler oluşur, dolaşımda sadece 8 birkaç gün kalırlar. Bağ dokuya geçerek makrofajları oluştururlar. Makrofajların belli başlı fonksiyonları şunlardır (10): • Fagositoz Makrofajlar bakteri, makromolekül, antijen gibi yabancı maddelerle, hasarlı, ölü hücre ve artıklarını fagosite ederler. Makrofajların hücre membranında değişik maddeler için spesifik reseptörler bulunur. Makrofajların yabancı partikülleri ve hasarlı dokuları tanımasında fosfolipid ve şeker reseptörlerinin rolü vardır. Bunlar içinde mannoz reseptörü, membran CD14 reseptörü, toll-like reseptör ailesi sayılabilir. Makrofajlar, antikorları ise yüzeylerindeki Fc reseptörleri ile tanırlar. Kompleman için de yüzeylerinde kompleman reseptörleri bulunur (10). • Salgılama Makrofajlar biyolojik olarak aktif olan pek çok madde salgılarlar. En önemlileri arasında TNF-α, interlökin-1β, IFN α, β, IL-6, 10, 12, fibroblast büyüme faktörü, prostaglandinler, kemokinler, nitrik oksit bulunmaktadır. Makrofajlar salgıladıkları tümör nekrozis faktör ile malign tümör hücrelerini öldürürler (10). • Antijen sunma İmmün regülasyonda antijenik uyarıyı, antijen sunan hücre olarak T hücrelerine arz ederek immün cevabın oluşmasına katkıda bulunur. Antijen sunma işlevinin ilk basamağı; antijen alımı, yıkımı ve MHC moleküllerine yüklenmesini içeren antijen sunumudur. Antijen sunumunun kapasitesi makrofajların molekülleri non-spesifik ve reseptör-aracılı fagosite etmeleriyle ilişkilidir. Antijen alımını izleyen dönemde antijenin proteolitik peptidlere işlenmesi ve MHC sınıf I ve sınıf II moleküllere yüklenmesi gerçekleşmektedir. Peptid-MHC kompleksi birlikteliğinde T hücre uyarımı sekonder lenfoid organların T hücre alanlarında gerçekleşmektedir. T hücreler peptidMHC kompleksine ko-stimülatör moleküllerin (CD80, CD86 gibi) varlığında T hücre reseptörü aracılığıyla bağlanırlar. T hücre aktivasyonuyla proliferasyon ve diferansiyasyon için gerekli olan yeni gen ekspresyonu indüklenmektedir. Son olarak CD4+ T hücre yanıtının fonksiyonel olarak polarizasyon aracılığıyla tip 1 ve tip 2 9 diferansiyasyon yolaklarına dönüşmesidir. Bu yanıtların gelişiminde antijen sunan hücrelerin salıverdikleri sitokinlerin rolü vardır (11). 2.1.3.Polimorfonüklear granülositler Polimorfonüklear granülositler veya lökositler (PMNL), kemik iliğinde dakikada yaklaşık 80 milyon hızda üretilmekte olup 2-3 günlük ömürleri vardır. İsminden de anlaşılacağı gibi olgun polimorflarda multilobüle çekirdek ve pek çok granüller bulunur. Bu granüllerin histolojik boyalarla boyanma karakteristiğine göre nötrofil, bazofil ve eozinofil olarak 3 ana grupta incelenirler (12). 2.1.3.1.Nötrofiller Nötrofiller, kandaki lökositlerin % 55-65’ini, dolaşımdaki polimorfların ise % 90’ını oluştururlar. Güçlü fagositik aktiviteleri vardır. Bunlar immün ve immün olmayan mekanizmalar ile aktive olabilir. Salgıladıkları sitokin ve granül materyalleri ile etki ederler (12). 2.1.3.2.Bazofiller ve Mast Hücreleri Bazofiller dolaşımda çok küçük miktarlarda bulunmaktadırlar. Büyük granüllerle dolu ve zayıf fagositik aktiviteye sahiptirler. Mast hücreleri ise dolaşımda bulunmazlar, dokularda; mukozal ve bağ doku olmak üzere iki çeşittirler. Bu hücrelerin fagositik aktiviteleri yoktur. Uyarıldıkları zaman histamin, prostaglandin ve lökotrien gibi mediyatörleri ortama verirler, IgE bağlayabilirler. Bu özelliklerinden dolayı anaflaktik allerjiden sorumludurlar (12). 2.1.3.3.Eozinofiller Eozinofiller, dolaşımdaki lökositlerin % 2-5’ini oluştururlar. Bu hücreler, mast hücreleri ve bazofillerden salınankemotaktik faktörler ile enflamasyon alanına doğru yönelirler. Fagositik aktiviteleri zayıftır. Allerjik ve paraziter olaylarda bölgesel ve sistemik sayıları artar (12). 10 2.1.4.Major Histokompatibilite Kompleksi İlk kez lökositlerde gösterilmiş olmalarından dolayı ‘İnsan Lökosit Antijenleri-Human Leukocyte Antigens (HLA)’ olarak adlandırılan, HLA antijenlerinin oluşması, organizmada ‘Majör Histokompatibilite Kompleksi-Major Histocompatibility Complex Gen Region (MHC)’ ismi verilen bir gen bölgesinin kontrolü altındadır. Üç ana gruba ayrılan bu bölgede MHC Sınıf-I (HLA-A, -B, -C, -E, -F, -G) MHC Sınıf-II (HLA-DR, DP, -DQ, -DO, -DN) ve MHC Sınıf-III (C2, C4A, C4B, PF, TNF-α,β) antijenleri bulunmaktadır (13). 2.1.4.1.Sınıf-I Molekülleri Sınıf-I molekülleri fizyolojik rollerine uygun olarak bütün çekirdekli hücrelerde mevcutturlar. Bir antijenin CD8 (killer) T lenfositlerce tanınabilmesi ancak Sınıf-I molekülü ile bir arada tutulmuş olmasına bağlıdır. Bu olgu HLA restriksiyonu olarak adlandırılır. Örnek olarak, bir virüs bir hücreyi enfekte ettiği zaman, viral antijenler peptid fragmanlarına ayrılırlar. Daha sonra Sınıf-I molekülleri tarafından bağlanarak CD8 killer T hücrelerine sunulurlar (14). Belirli bir T lenfositin antijen reseptörü, belirli bir viral peptidi ancak belirli bir Sınıf-I molekülü varlığında tanıyabilir. Bu reseptörler farklı Sınıf-I molekülü ile bağlı özgün bir peptidi özgün bir Sınıf-I molekülü ile bağlamış farklı bir viral peptidi veya bağlı olmayan bir Sınıf-I molekülünü tanımazlar. Tanıma gerçekleştiğinde killer T lenfositler (CD8) viral antijen taşıyan hedef hücreyi öldürürler (14). 2.1.4.2.Sınıf-II Molekülleri Sınıf-II MHC moleküllerinin ana işlevi CD4 T-lenfositlere ekzojen kaynaklardan elde edilmiş işlenmiş antijen sunmaktır. Böylece MHC Sınıf-II molekülleri antijen spesifik immün yanıtın başlatılması için kritik öneme sahiptir. Antijen sunumu yanında giderek artan kanıtlar gösteriyor ki MHC Sınıf-II molekülleri ayrıca sık apoptozise götüren intraselüler sinyal yolaklarını aktive etmektedirler (15). 11 2.1.4.3.Sınıf-III Molekülleri Sınıf–III molekülleri immün fonksiyonlarla salınan çeşitli proteinlerdir. Sınıf-III genlerinin Sınıf-I ve Sınıf-II genleri ile aynı fonksiyonları paylaşmadığı açıktır. MHC Sınıf-III antijenleri Sınıf-I ve Sınıf-II antijenleri arasındaki bölgede bulunurlar. Klasik kompleman yolunun C2, C4a ve C4b, alternatif yolun properdin faktörü ile 21hidroksilaz gibi enzimler ile TNF gibi sitokinlerin sentez edildiği bölgedir (17). 2.1.5.Kompleman Sistemi (Tamamlayıcı Sistem) Doğal bağışıklık sistemi enfeksiyon ajanlarına karşı ani immün cevapta çok önemli rol oynar. Kompleman sistemi, doğal immün sistemin bir parçası olarak sonradan kazanılmış bağışıklıkla birlikte patojenlerin öldürülmesi ve immün komplekslerin temizlenmesinde görevlidir (16). Tanımlanan tamamlayıcı bileşenleri düzenlemek için, her faktöre keşfedilme sırasına göre bir numara atanmıştır; örneğin; C1, C2, C3 ve C4 gibi. Daha sonraki çalışmalarda bu faktörlerin reaksiyon verme sırasının, keşfedilme sırasıyla aynı olmadığı görülmüştür. 1930’lu yılların sonlarına doğru ilk dört bileşenin reaksiyon sırası şu şekilde değiştirilmiştir. C1, C4, C2, C3. Bu tamamlayıcı zincir, insanları patojenik mikroorganizmaların saldırılarından koruyan güvenlik ağının bir parçası olarak düşünülmüştür. Enfeksiyonla ilişkili bir uyarıcı, kompleman yanıtın başlangıç noktası olarak kabul edilmiştir. Koruyucu yanıt, bulaşıcı hastalık perspektifinden oluşturulduğu için, C1-C4-C2-C3 dizisi, kompleman sisteminin klasik yol (classical pathway) girişi olarak adlandırılmıştır (16). Daha sonraki çalışmalar bu klasik yolun dışında bir etki mekanizması daha olduğunu göstermiştir. Bu çalışmalar, alternatif yolu (alternative pathway) tanımlayacak öncü çalışmalar idi. Bu çalışmalardan bir diğeri de Dr. Louis Pillemer’in daha önceden isimlendirilememiş olan serum proteini ‘properdin’ i tanımlamasıdır. Bu protein antikor varlığına gereksinim duymadan komplemanı aktive ederek bakteri ve mayaya bağlanmaktadır (16). 12 Şekil 2.2. Klasik ve alternatif kompleman yollarının aktivasyonları ve membran atak kompleksi (C5b-9) oluşumunun basamakları (17). Komplemanın başlıca biyolojik aktiviteleri şunlardır; fagositozu güçlendirmek, anafilatoksik etki, kemotaksis, viral nötralizasyon, sitoliz, immün modülasyonu (12). 13 2.1.6.Sitokinler Sitokinler; lenfositler, iltihabi hücreler ile endotelyal hücreler gibi immün sistem hücreleri arasındaki etkileşmeleri düzenleyerek aktivitelerini yönlendiren polipeptit yapısında kısa etkili ve çözünür moleküllerdir (18). Salgılandıkları zaman organizmada sistemik (endokrin) ve/veya salgılandıkları hücre etrafındaki hücrelere (parakrin) veya doğrudan salgılandıkları hücreler üzerine (otokrin) etkide aracılık ederler (12). Lenfosit kaynaklı sitokinler lenfokin, makrofaj/monosit kaynaklı sitokinler monokin, lökositlere etki eden monokin ve lenfokinler ise interlökin (IL) diye adlandırılırlar (18). Kemokin ise kemotaktik ve sitokin parçalarının birleştirilmesiyle üretilen makrofaj ve monositleri enfeksiyon bölgesine çekebilen bir grup sitokindir (12). Görevleri şunlardır: • Lenfoid hücrelerinin aktivasyonunu, çoğalmasını ve farklılaşmasını sağlamak (spesifik immünite) • İmmün cevabı şiddetlendirerek veya baskılayarak regüle etmek • Enflamasyon olaylarına katılan hücreleri aktive etmek, olay yerine toplayarak orada tutmak (non-spesifik immünite) • Kemik iliğine etki ederek hemopoietik regülasyona katılmak • Bazı hipofiz hormonlarının ve diğer biyolojik maddelerin sentez ve salınmalarına sebep olmak (12). Bazı önemli sitokinler: • İnterlökin 1 Esas olarak makrofajlar tarafından üretilen ve pek çok hedef hücrenin farklılaşmasını ve özgül ürünler sentezlemesini sağlaması yanında T hücrelerini farklılaşma ve IL-2 üretme yönünden aktif hale getiren bir proteindir. Endojen pirojen olup santral sinir sistemi üzerine IL-1’in etkisi ile ateş ortaya çıkar. Prostaglandin sentezi gibi enflamatuar yanıtları indükler, lökositlerin endotel hücrelerinde adezyonunu sağlar ve vasküler geçirgenliği artırarak PMNL ve makrofajların bölgeye göçüne neden olur (12). 14 • İnterlökin 2 T hücresi büyüme faktörü de denilen (TCGF) IL-2, T lenfositlerinin hücre siklusunun G1 fazından S fazına ilerlemesinden sorumlu olan sitokindir. IL-2, CD4+ T hücreleri tarafından üretilir az olarak da CD8+ T hücreleri tarafından üretilir. IL-2, kendisini üreten hücrelere etki edip kendi oluşumunu sağlar, bu onun otokrin büyüme faktörü görevini gösterir. Ayrıca parakrin büyüme faktörü olarak da etkisi bulunur. IL-2, T lenfositleri için otokrin büyüme faktörüdür ve IFN-γ, lenfotoksin gibi T hücresinden kökenini alan sitokinlerin sentezini uyarır. NK hücrelerinin büyümesini uyarır ve onların sitolitik işlevlerini artırır. Bunu lenfokinle aktive edilmiş öldürücü hücreler (LAK) üreterek yapar. Ayrıca IL-2, insan B lenfositlerine etki ederek hem büyüme faktörünün hem de antikor sentezi uyaranı olarak etki gösterir (19). • İnterlökin 4 ve 5 CD4+ T lenfositleri tarafından üretilirler ve B hücrelerinin üreme ve farklılaşması üzerine etkilidirler. IL-4 ayrıca MHC Sınıf II ekspresyonunu atırır ve IL-3 ile birlikte mast hücrelerinin gelişiminde sinerjik rol oynar (8,12). • İnterlökin 6 Başlıca makrofajlar daha az olarak da B, T, endotel hücreleri ve fibroblastlar tarafından üretilir. B hücreleri üzerinde gelişmeyi sağlayıcı etki gösterir. IL-1 ile birlikte bir kofaktör olarak olarak IgM sentezletir, IL-5 ile birlikte IgA sentezini uyarır (20). • İnterlökin 8 IL-8’in hedef hücreleri ise nötrofiller ve T hücreleridir. Nötrofillerin mobilizasyonunu, aktivasyonunu ve degranülasyonunu sağlar, angiogenezde rolü vardır. IL-8 lökositlerin vasküler endotele stabil olarak bağlanması için adezyonunda rol oynar. İkincil olarak infeksiyon alanında konsantrasyon artışı vasıtasıyla nötrofillerin migrasyonunda gradyan sağlar (21). 15 • İnterlökin 10 İnterlökin-10 insan immün cevabında bulunan en önemli antiinflamatuar sitokindir. Primer olarak T lenfositleri, monositler, makrofajlar, B lenfositleri ve nötrofiller tarafından sentezlenirler. IL-10 koruyucu aktivitesini IL-1β, TNF-α, IL-8, IFN γ, IL-6 ve prostaglandin metabolitleri gibi inflamasyon mediyatörlerini inhibe ederek gösterir. Çeşitli tümörlerde seviyesinin arttığı görülmüştür (22). • İnterlökin 12 IL-12 hem doğal hem de kazanılmış bağışıklıkta önemli rol oynayan bir sitokindir. Özellikle B hücreleri, dendritik hücreler ve makrofajlar olmak üzere birçok farklı hücre tarafından üretilmekte olup, T ve NK hücrelerinin sitotoksisitesini artırmakta, başta IFN-γ, TNF-α, GM-CSF, M-CSF, IL-3, IL-8 olmak üzere birçok sitokin ile kemokinin salgılanmasını uyarmaktadır (23). • Tümör nekroz faktörü Esas olarak makrofajlar (TNF-α) ve T lenfositler (TNF-β) tarafından sentezlenerek nötrofillerin fagositoz ve öldürme etkinliklerini aktifleştirir (12). • Granülosit makrofaj-koloni stimüle edici faktör GM-CSF, aktive T hücreleri, endotelyal hücreler, fibroblastlar ve makrofajlar tarafından sentez edilen, glikoprotein yapısında bir büyüme faktördür (24). PMNL ve eozinofillerde myeloproliferasyon, PMNL ve makrofaj kemotaksisi ve makrofajlardaki sitokin sentezini artırmaktadır. GM-CSF’nin ayrıca apoptozise karşı koruma, reaktif oksijen üretim artışı ve nötrofillerde bakterisidal aktivite artışı yapması gibi rolleri de vardır (24). 16 Şekil 2.3. GM-CSF ve T hücre yanıtları (24) • TGF-β (Değişiklik yapan büyüme faktörü-β) Trombositler, T ve B lenfositler tarafından üretilir, B hücre proliferasyonunu engelleyici etkileri vardır. IgA üretiminde ise uyarıcı etkisi vardır (20). • İnterferonlar İnterferonlar, geniş biyolojik etkiye sahip, birçok indükleyicilerin varlığında salgılanan son derece etkili sitokinlerdir. Yapısal, biyokimyasal ve antijenik özellikleriyle üç alt gruba ayrılırlar. IFN-α monositler, TNF-α, IL-1 ve virüslerin uyarısıyla üretilmektedir. IFN-β fibroblastlardan ve epitelyal hücrelerden IFN-γ ise CD4+ ve CD8+ T hücreleri, NK hücreleri ve makrofajlarda üretilmektedir. İnterferonlar çeşitli hücrelerin yüzeylerindeki reseptörlere bağlanarak etkilerini gösterirler. Reseptöre bağlanan IFN molekülü, hücrede antiviral, antiproliferatif ve immünomodülatör etki göstermektedir (25). 17 Şekil 2.4. İmmün sistemin doğuştan ve kazanılmış bölümlerinin ana bileşenlerinin ve hedeflerinin şematik özeti (9) 2.2. TÜMÖR İMMÜNOLOJİSİ Son yıllarda immünoloji alanında elde edilen gelişmeler, organizma ve tümör etkileşimine de yansımıştır. Bilinen immünolojik mekanizmalar çerçevesinde tümör immünolojisi üç ana başlık altında incelenebilir: Tümör antijenleri, efektör mekanizmalar ve kanserin immüniteden kaçış mekanizmaları (26). 18 2.2.1. Tümör Antijenleri Yapılan çalışmalar tümör hücresinde bazı antijenlerin varlığını ortaya koymuştur. Üç gruba ayrılır. Tümör assosiye antijenler: Kanser hücrelerinde olduğu gibi normal doku hücrelerinde de bulunabilen antijenler bu grupta incelenir. Tümör spesifik antijenler: Bu grup sadece tümör hücresine özgüdür, normal hücrede bulunmaz.Tümör spesifik transplantasyon antijenleri: Konakta immün cevabı uyaran, tümör büyümesine karşı direncin ve tümör rejeksiyonuyla sonuçlanan bir dizi reaksiyonun oluşmasına sebep olan antijenlerdir (26). 2.2.2. Efektör Mekanizmalar • Sitotoksik T-lenfositleri (CTL) CTL-tümör hücresi etkileşiminde ilk adım, hedef hücredeki tümör antijeninin kendisine özgün CTL-reseptörü tarafından tanınmasıdır. Bu tanınmanın gerçekleşmesi ve bundan sonraki sitolitik etkinin ortaya çıkabilmesi için tümör antijeninin tümör hücresinin sınıfI MHC antijeni ile algılanması gerekmektedir. Aynı olay yardımcı T hücrelerinde de benzer biçimdedir fakat sınıf-II MHC ile gerçekleşmektedir. Buna MHC ile sınırlı tanıma (MHC-Restricted recognition) denmektedir. Hedef hücreyle temasa geçen CTL polarizasyon gösterir. CTL sitoplazmasında bulunan sitolitik granüller temas yerinde toplanır. Hedef hücre tamamen granüllerler kaplanır ve membranında oluşan delikler nedeniyle lizise uğrar. CTL granülleri aynı zamanda hedef hücre DNA’sını da tahrip edebilir (26). • Makrofajlar Makrofajların tümör immünitesinde önemli bir rolü vardır. Antijen sunucu hücreler olarak görev yaparak tümör hücrelerinin lizisinde rol alan efektör mekanizmaları uyarırlar. Makrofajlar MHC sınıf II molekülleri ile tümör antijenlerini yardımcı T hücrelere sunarlar. Makrofajlar, yardımcı T hücrelerinden salınan INF-γ, granülositmakrofaj koloni uyarıcı faktör (GM-CSF), TNF, IL-4 gibi makrofaj aktive edici faktörler (MAF) ile aktive olur. TNF, NO, O2, proteazlar gibi inflamatuvar mediyatörlerin ortama salınmasıyla tümör hücreleri sitolitik olarak makrofajlarca elimine edilirler. Ayrıca makrofajlar, Fc reseptörleri veya kompleman reseptörleri ile 19 antikorlar tarafından bağlanmış tümör hücrelerini yakalayarak ‘antikor bağımlı hücre aracılı sitotoksisite (Antibody dependent cell mediated cytotoxicity=ADCC) mekanizmasıyla da öldürürler (27). • NK hücreler NK hücreleri birçok tümör hücresini hedefleyerek yok eden hücrelerdir. NK hücreleri tümör hücrelerini sitolitik olarak öldürürler. NK hücreleri tümör hücrelerinin MHC moleküllerine bağlanmazlar. NK hücreleri MHC sınıf I molekülleri olmayan veya bu molekülü çok az miktarda bulunduran tümör hücrelerini tanırlar. Tümör hücre yüzeyindeki NK reseptörlerine bağlanarak aktive olurlar. Tümör hücresinde deliklere ve hasara yol açarlar. Antikorlarla çevrili tümör hücrelerini Fc reseptörleri ile tanıyarak ADCC ile tümörleri yok ederler (27). • LAK hücreleri İnterlökin 2 adlı lenfokinle aktive edilen lenfositler, tümör hücrelerini lizise uğratabilme yeteneğini kazanırlar. Bunlara lenfokince aktive edilmiş öldürücü hücreler anlamına gelen LAK adı verilir (26). • B lenfositler Tümör lizisinin antikora bağımlı yoldan gerçekleştirilmesinde iki mekanizma görev yapar; i) Komplemana bağımlı sitotoksisite ve ii) Antikora bağımlı sitotoksisite. Birincisinde tümör hücresinin yüzeyinde bulunan Fc reseptörlerine komplemanı fikse eden antikorlar bağlanır. Kompleman aktivasyonuyla tümör hücre membranında ‘Membran Atak Kompleksi (MAC)’ oluşur. Delikler açılır ve lizis olur. Alternatif mekanizma olan ADCC ile NK hücreler, makrofajlar, granülositler gibi fagositer hücreler hücre lizisine aracılık ederler. Anti-tümör antikorlarla kaplanmış tümör hücreleri Fc bölgelerinden Fc reseptörleri bulunduran NK hücrelerine tutunurlar. NK hücreleri tümör hücrelerini lizis ile yok ederler. ADCC mekanizması kompleman bağımlı sitotoksisiteye kıyasla tümör eliminasyonunda daha etkili bir mekanizmadır (27). 20 2.2.3. Kanserin İmmün Sistemden Kaçış Mekanizmaları Kanser hücresinin immün efektör mekanizmalardan kaçmasını sağlayacak çeşitli yollar bulunmaktadır. • Tümör hücresi membranındaki antijenik değişimler: Tümör hücresi antijenleri immün sistemi uyaracak miktarda sentez edilmez veya çok kısa sürede modülasyon göstererek immüniteden kaçar. • Antijen-antikor kompeksleri: Dolaşımdaki antijenler, antikorlar ile kompleksler oluşturarak immün yanıtın ortaya çıkmasını engelleyebilirler. Bu antijen-antikor kompleksleri katil hücrelerin Fc reseptörlerine bağlanarak onları bloke ederler. Böylece antikora bağımlı hücresel sitotoksisite reaksiyonlarını engellerler. • Spesifik ve non-spesifik supresör hücreler: Tümör assosiye antijenlerin bazıları spesifik supresör T-lenfosit gruplarının oluşmasına yol açmaktadır. • Tümör tarafından oluşturulan supresyon: Bazı tümörler salgıladıkları prostaglandinler aracılığı ile immün yanıtı baskılayabilmektedirler. Bazen tümör hücrelerince lenfoit dokunun istila edilmesi, immün yanıtı gerçekleştirecek hücrelerin azalması ile sonuçlanmaktadır. • Diğer faktörler: İmmün sistemi etkileyen bazı genetik ve edinsel hastalıklar sonucu maligniteye eğilim artmaktadır. Örneğin yardımcı T-hücre sayısının azaldığı AIDS hastalığında malignite oranı yüksektir (26). 2.3.ANTİKORLAR Antikorlar kendi ağır zincir bölgesinin dizisini esas alan beş sınıfa ayrılır: IgM, IgD, IgE, IgA ve IgG. IgG, kanser immünoterapisinde en sık kullanılanlardır. Antikorlar, antijen bağlayan parçası (Fab) ve sabit parçası (Fc) olmak üzere iki ayrı işlevsel bölgeye ayrılırlar (28). Fab, antikorun antijen bağlama bölgesini oluşturan ve antijen spesifitesini sunan tamamlayıcılığı belirleyen çok değişken üç bölgeyi içerir. Antikorların immün efektör fonksiyonları, kompleman-bağımlı sitotoksisiteyi (CDC) başlatma yeteneğinde olan, Fc 21 γ-reseptörlerine (FcγR) ve neonatal Fc reseptörlerine (FcRn) bağlanan Fc parçası ile bağlantılıdır (28). Şekil 2.5. Antikorun yapısı (29) Ig’nin alt sınıflarından olan IgG1 ve IgG3, klasik kompleman yolunun güçlü aktivatörlerindendir. Hücre yüzeyine iki ya da daha fazla IgG bağlanması, C1r’nin enzimatik aktivitesinin ve daha sonraki kompleman proteinlerinin aktivasyonunu takiben C1q’nun Fc parçasına yüksek afiniteli bağlanmasına yol açar. Bu aşamanın sonucu, MAC tarafından tümör hücre lizislerinde ve tümör hücre yüzeyinde porların oluşturulmasıdır. Buna ek olarak, son derece kemotaktik kompleman molekülleri C3a ve C5a’nın üretimi makrofaj, nötrofil, bazofil, mast hücreleri ve eozinofil gibi immün efektör hücrelerinin alımına ve aktivasyonuna yol açar (28). Fcγ reseptörleri, immünoreseptör tirozin bazlı aktivasyon motifleri (ITAMs) aracılığıyla aktivasyon sinyallerini aktarabilir veya immünoreseptör tirozin bazlı inhibitör motifleri (ITIMs) aracılığıyla inhibe edici sinyal sağlayabilir. Majör inhibitör Fcγ reseptörü tek zincir FcγRIIB (CD32 olarak bilinen)’dir. Oysa Fc-bağımlı uyarıcı sinyaller FcγRI (CD64 olarak bilinen) ve FcγRIIIA (CD16A olarak bilinen) tarafından aktarılır, her ikisi 22 de sinyal aktarımını başlatmak için ITAM içeren γ zinciri gerektirir. FcγRI makrofajlar, dendritik hücreler, nötrofiller ve eozinofiller tarafından eksprese edilen yüksek afiniteli bir reseptördür. FcγRIIIA NK’lar, dendritik hücreler, makrofajlar ve mast hücreleri tarafından eksprese edilen primer aktive edici Fcγ reseptörüdür (28). Tümör hücrelerine IgG antikorlarının bağlanması, Fcγ reseptörlerini ekprese eden NK’lar, nötrofiller, mononükleer fagositler ve dendritik hücreler gibi immün efektör populasyon tarafından hedeflerin tanınmasını sağlar. Bu hücreler üzerinde Fcγ reseptörlerinin çapraz bağlanması tümör hücre tahribatını arttırır. Tümör hücresinin lizisini takiben antijen sunan hücreler MHC Sınıf II molekülleri üzerindeki tümör kaynaklı peptitleri sunabilir ve CD4+ T hücrelerin aktivasyonunu artırabilir. Buna ek olarak, çapraz-sunum olarak bilinen CD8+ T hücrelerinin aktivasyonuyla sonuçlanan bir işlemde, tümör kaynaklı peptitler MHC sınıf I moleküllerine sunulabilirler (28). 2.3.1. Monoklonal Antikor Monoklonal antikorlar hibrit hücreler tarafından üretilen homojen antikor topluluklarıdır. Bu antikorlar, bağışıklanmış fare veya ratlardan elde edilen dalak hücreleri ile myeloma adı verilen lenfoit tümör hücrelerinden geliştirilen melez hücreler tarafından üretilirler. Her iki tip hücrenin özelliklerini taşıyan hibridomalar, doku kültürlerinde çok miktarda mAb oluştururlar (30). 1975’te Georges Köhler ve Cèsar Milstein Nature’de hibridoma teknolojisi tarafından üretilen bir işlem yayınlamışlardır. Bu işlem, B hücresi tarafından üretilen spesifik bir antikor ile bir myeloma hücresi arasındaki füzyondan çıkan hibrit hücre hatlarıdır. Bunu yaparken, Köhler ve Milstein aynı özellikte, monoklonal denilen çok miktarda antikor üreten bir yöntem sağlamışlardır (31). 1984 yılında, bu atılım buluş sayesinde, Köhler ve Milstein, immünolojiye olan katkıları sebebiyle Niels Jerne ile tıp ve fizyoloji dalında Nobel ödülü almışlardır (31). Monoklonal antikorların üretiminde rekombinant DNA teknolojisi ve hibridoma teknolojisi kullanılmaktadır. Rekombinant DNA teknolojisi, genetik rekombinasyon olaylarının yapay olarak gerçekleştirilmesi esasına dayanan ve istenilen herhangi bir gen ya da ürünün çoğaltılmasını mümkün kılan bir yöntemdir. Rekombinant DNA 23 teknolojisi, bugün dev adımlarla ilerlemekte olan bir gen mühendisliği konusudur. Bu teknoloji ile çeşitli amaçlar için adeta ısmarlama genler ve proteinlerin üretimi mümkün olmaktadır (32). Rekombinant DNA teknolojisinde izlenen basamaklar kısaca şöyledir: Çoğaltılması istenen gen önce orijinal kromozomdan restriksiyon endonükleaz enzimi yardımıyla kesilerek alınır ve plazmid ya da faj gibi bir vektöre integre edilir. Sonra bu vektör transformasyon yoluyla bir bakteri ya da maya içine sokulur ve daha sonra bu mikroorganizmaların kültürü yapılarak istenen gen veya protein gerekli miktarlarda elde edilir (32). Bir antijen (bakteri, virüs, hücre veya molekül) ‘epitop’ adı verilen birçok antijenik bölge içerir. Her epitop ayrı bir antikor tipi tarafından tanınır. Bir antijen organizmaya girdiğinde her epitop için bir grup B-lenfosit hücresi özgül antikor üretmeye ve bölünerek çoğalmaya başlar. Böylece her epitop için özgül antikor üreten bir B-lenfosit kolonisi oluşur. Bir antijene karşı değişik B-lenfosit kolonileri oluştuğu için elde edilen antikora ‘poliklonal’ (çok kolonili) antikor adı verilir. Köhler ve Milstein’ın geliştirdikleri teknik, bir tek epitopa özgü (monoklonal) antikorların, hem de sınırsız miktarlarda elde edilebilmelerine olanak sağlayarak, moleküler biyolojiye önemli bir hız kazandırmış durumdadır (33). Bu hibridoma tekniğinin temelinde üç bilgi bulunur: • B-lenfositler tek bir epitopa özgü antikor üretip salgılayan, yaşam süreleri birkaç günle sınırlı kan hücreleridir. • Tümör hücreleri bölünerek çoğalma kontrolünü kaybetmiş, hızla üreyen ölümsüz hücrelerdir. • Belli koşullarda aynı organizmaya ait değişik hücreler birleştirilerek her iki hücrenin özelliklerini taşıyabilen melez hücreler (hibridoma) elde edilebilir (33). Monoklonal antikorlar, antijen üzerinde bulunan ve epitop denilen antijenik determinantlardan bir tanesine hedeflendikleri için enfeksiyon hastalıklarının veya tümörlerin tanı ve tedavisinde etkin olarak kullanılırlar. Hibridoma teknolojisi ile fare kaynaklı olan ilk mAb’lardan sonra rekombinant DNA teknolojisi ile elde edilen insan kaynaklı veya bir kısmı insana özgü bölgeler içeren tipleri üretilmektedir (34). 24 Sağlıklı bir insanın serumundan fraksiyonlanan IgG’nin elektroforetik tekniklerle ayrıştırılması sonucunda, 105-106 farklı IgG molekülünün var olduğu bulunmuştur. Buna rağmen, miyolemal hastalıklardan muzdarip olan sağlıksız bir bireyin serumundan elde edilen IgG fraksiyonu ise çok daha az sayıda IgG molekülleri içermektedir ve bu moleküllerin büyük bir çoğunluğunu ise tek bir tip IgG molekülü oluşturmaktadır. Böyle bireylerin dolaşım sistemlerinde sirküle eden lenfositlerin çoğu, miyolema hücreleridir ki bunlar, neoplastik B lenfositlerinin tek bir klonunu içermektedirler (35). Monoklonal antikor üretimi fare ve ratların B-lenfositi myeloma tümörlerinden elde edilen hücre hatlarının geliştirilmesi ile mümkün olmuştur. Myeloma hücreleri kendi çeşitli Ig’lerini sentezleme yeteneğini kaybetmişlerdir. Monoklonal antikor üretiminde, fare ve rat myeloma hücreleri kullanılmaktadır. Rat myeloma hücreleri, daha büyük olmaları ve daha kolay kaynaşmaları nedeniyle fare myeloma hücrelerine tercih edilirler (30). Normal B lenfositlerine benzemeyen miyolema hücreleri ise hızlı bir şekilde bölünmeye devam eder ve hiçbir antijenin stimüle etkisi bulunmadan tek bir tip Ig molekülünü salgılarlar. Normal B lenfositleri in vitro koşullarda birkaç saatten fazla canlı kalamamalarına rağmen, miyolemalar diğer neoplastik hücreler gibi hücre kültür ortamlarında ölümsüz (immortal) olarak kalabilirler (35). Normal memeli hücreleri iki farklı yoldan DNA moleküllerini sentezlerler: • De novo (yeni baştan sentez) metabolik yolu ile nükleik asitler, pürin ve pirimidin bazlarından, deoksiriboz ve inorganik fosfattan oluşturulurlar. • Kurtarma metabolik yolunda ise, benzer nükleotidlerin dönüştürülmesi ile doğru nükleik asitlerin sentezi sağlanır. De novo metabolik yolu aminopterin ile tamamen inhibe olmaktadır. HGPRT enzimi (hipoksantin-guanin fosforiboziltransferaz) içeren hücreler ise kurtarma metabolik yolunu kullanarak, aminopterin içeren besi ortamlarında canlı kalabilirler. Buna rağmen, kurtarma metabolik yolunun işletilerek canlılığın sürdürülebilmesi için ortama hipoksantin ve timidin eklenmek zorundadır. Bu maddelerin eklendiği besiyeri ise HAT besiyerleri (hipoksantin, aminopterin, timidin) olarak adlandırılırlar. Böylece HAT besiyeri, HGPRT enzimi içeren hücrelerin, bu besiyerinde seçici olarak büyümelerini sağlarken; HGPRT içermeyen (HGPRT-) hücrelerin büyümelerini engelleyecektir. Fare 25 miyolema hücrelerinin HGPRT- olan klonları seçilir ve bunlar HAT besiyerinde canlılıklarını sürdüremezler ve çoğalamazlar. HGPRT- olan miyolema klonları seçilirken bunların, istenmeyen Ig’leri çok az miktarda veya hiç üretmeyenleri seçilmelidir. Günümüzde, mAb üretiminde kullanılan standart metot ise Şekil 6’da verilmiştir. Uygun bir antijen ile aşılanmış olan farenin dalak hücreleri ise, duyarlı hale getirilmiş olan B lenfositlerinin süspansiyon kaynağı olarak kullanılır. In vitro koşullarda büyüyen, duyarlı B lenfosit hücreleri ve HGPRT üreten miyolema hücreleri, füzyon oluşturmak amacıyla, hücrelere toksik olan %30-50 (w/v) polietilenglikol (PEG) içeren ortamda birkaç dakika süre ile karıştırılır (35). Standart bir füzyon için ortamda 1 x 108 lenfosit ve 2 x 107 myeloma hücresinin bulunması gereklidir. Hücreler mikrotitrasyon kapları içindeki kültür vasatında 37˚C’de inkübe edilir.Daha sonra, bu füzyon karışımı ise HAT besiyeri içeren kültür ortamlarına aktarılır (30). Lenfositler ve füzyon oluşturan lenfositler, in vitro koşullarda kültürlerinin yapılamamasından dolayı çok kısa bir sürede ölürler. Miyolema hücreleri ve füzyon oluşturmuş olan hücreleri ise HGPRT içermedikleri için HAT besiyerinde bulunan aminopterin tarafından zehirleneceklerdir. Sadece B lenfositleri ile HGPRT pozitif olan miyolema hücrelerinin füzyonları sonucunda oluşan hibridomalar bu ortamda canlı kalacaklardır. Çünkü, miyolema hücreleri, hibrit hücrelere in vitro koşullarda yaşama özelliğine aktarırken, B lenfositleri ise bu hibrit hücrelerin HAT besiyerlerinde üreyebilmeleri için gerekli olan HGPRT enzimini aktaracaklardır. 10-14 gün sonra ise kültür ortamında canlı kalan hücreler sadece hibridomalardan ibaret olacaklardır. Buna rağmen, bu hibridomaların birçoğu ya istenmeyen Ig’leri üretebilir ya da hiçbir Ig üretmeyebilir. Ne yazık ki, bu hibridomalardan hiç Ig üretmeyenler, Ig üretenlere oranla çok hızlı büyümektedirler. Bu nedenle, hibridoma oluşumunu takip eden 7-14 gün boyunca, her bir kültürün devamlı olarak istenilen Ig’yi üretip üretmediğinin test edilmesi ve üretmeyenlerin elimine edilmesi gerekmektedir (35). Füzyon işleminden sonra oluşan hibridlerin tümünün antikor sentezleme yeteneğinde olup olmadıkları bilinmemektedir. Bu nedenle, kullanılan antijene karşı hibridomalar tarafından üretilen antikorlar, hücrelerin ürediği besiyerinin üst sıvısında aranırlar. Füzyon işlemi rastgele olduğundan, antikor sentezleyen hibridlerin oranı % 1-50 arasında değişir. Antikorların üst sıvıdaki varlıklarının 2-3 gün gibi kısa bir sürede 26 kontrol edilmesi iki yönden önem taşımaktadır. Mikrotitrasyon kabının çukurlarında hızla üreyen melez hücreler metabolizma artıklarının çoğalması nedeniyle ölebilirler. Diğer yönden, antikor üretmeyen hücreler hızla çoğalarak, antikor üretenleri baskılayabilirler. Üst sıvıdaki antikorların belirlenmesi amacıyla kullanılan antijenin özelliğine göre radyoimmunoassay, ELİSA ve hemolitik plak deneyi gibi duyarlı testlerden yararlanılmaktadır (30). Bunun yanı sıra, istenen ya da kullanışlı Ig üretenlerin ise alt-kültürleri hazırlanmalı ve proteinleri sıvı-azot içerisinde dondurularak saklanmalıdır. Aksi halde kültürlerin mikroorganizmalar ile kontaminasyonu sonucu çalışma başarısız olur (35). 27 Şekil 2.6. Monoklonal antikorların üretimi. (a) Monoklonal antikor üretiminde en yaygın olarak kullanılan prosedür. (b) İlgilenilmekte olan özgül hibridomaların oluşum oranını artırmak için, B hücrelerinin antijen-özgül adezyon ile miyolema hücrelerine bağlanmasını sağlayan prosedür (35) Günümüzde mAb’ların uygulamaları fazlaca yaygınlaşmıştır. Araştırma amaçlı olarak in vitro ve gittikçe artan bir hızla in vivo teşhis ve tedavi sistemlerinde kullanılmaktadırlar. 1998’de Amerika’da geliştirilen 350 tane biyolojik maddeden 74’ünün mAb oluşu, biyofarmasötik ürünler arasında mAb’ların önemini 28 göstermektedir. Kanser, transplantasyon ve bulaşıcı hastalıklar gibi türlü hastalıkların belirlenmesinde ve tedavisinde antikorlardan yararlanılmaktadır. Rekombinant teknolojisinin gelişmesiyle antikor kullanım alanı oldukça artmıştır. Bugün, faydalı antikor fragmentlerini üretmek için antikorları ‘insanlaştırmak’ ve istenen bölgeye hedeflemek için, antikorları izotop ya da ilaçlara bağlamak mümkündür. Son yıllardaki güzel bir gelişme de uygunluk onayı almış monoklonallerin sayısıdır (36). 2.3.2. Monoklonal Antikorlar ile Kanser Tedavisi Geleneksel antikanser terapötikleri zayıf terapötik indekse sahip olan, normal sağlıklı dokulara toksisite ile sonuçlanan ve spesifiteden yoksun terapötiklerdir (37). Monoklonal antikorlar geleneksel tedavinin aksine antikor aracılığıyla tümör antijenlerine hedeflendikleri için hem spesifiktirler hem normal dokulara toksisiteleri görece çok daha azdır hem de geliştirilen mekanizmalarıyla indeksleri daha fazladır. Tablo 2.1. mAb veya mAb ile ilişkili terapötikler için uluslararası son ekler (31) Son ek Molekülün tipi -aksomab Bispesifik fare-rat hibridi mAb -sept Fc füzyon proteini -omab Fare IgG2 -stim Fc füzyon peptidi -umab Faj görüntüleme veya transgenik fare teknolojisi insan antikoru -ksimab Kimerik IgG1 -zumab Humanize IgG1 Kanser tedavisiyle ilişkili en az altı sınıf mAb vardır. Birincisi, tümörün hücresel otonom yaşamsal basamaklarını inhice eden mAb’lardır. (Epidermal büyüme faktörü reseptörünü, EGFR, inhibe eden ve kolorektal kanser tedavisinde onaylı olan setuksimab ve panitumumab) (31) 29 Şekil 2.7. Panitumumab’ın etki mekanizması (38) Şekil 2.7’de: Panitumumab epidermal büyüme faktörü reseptörünü bloke ederek epidermal büyüme faktörünün ve diğer bağlanma faktörlerinin bağlanmasını önler. Epidermal büyüme faktörünün aktivasyonu ve sinyal aktarımının daha sonraki aşamaları inhibe edilerek büyüme faktörlerinin ekspresyonu azaltılır. Setuksimab ve erlotinib de ayrıca epidermal büyüme faktörü reseptörününün aktivasyonunu önler (38). İkincisi, tümör-stroma reaksiyonlarını engelleyen mAb’lardır. (Vasküler endotelyal büyüme faktörünü, VEGF, bloke eden ve akciğer, meme, böbrek, kolorektal kanser tedavilerinde kullanılan bevasizumab) (31) 30 Şekil 2.8: Tümör hücreleri, stroma hücreleri, perisitler ve endotel hücreleri arasındaki intraselüler sinyalizasyon (39) Şekil 2.8’de: Tümör/mikrovasküler mikroortamındaki selüler oyunculardır. VEGF, A1 ve Bcl-2 gibi antiapoptotik proteinlerin ekspresyonunu indükleyerek, fosfoinositid 3kinaz/Akt sinyal yolunu aktive ederek, nitröz oksidi ve prostasiklini uyararak, fokal adezyon kinaz tirozin fosforilasyonu artırarak böyle çeşitli yollar vasıtasıyla endotel hücreleri için antiapoptotik rol oynar. VEGF reseptör aktivitesi öncelikle ilgili ligandların varlığıyla düzenlenirler. Ligand ekspresyon düzeyleri hipoksi, çevresel stres ve glikoz eksikliği gibi birçok faktöre bağlıdır. EGF, endotelyal büyüme faktörü; HGF, hepatositik büyüme faktörü; PDGF, platelet kaynaklı büyüme faktörü; TGF, transforming growth factor; VEGF, vasküler endotelyal büyüme faktörü (39). 31 Üçüncüsü, eksprese antijenleri tümör hücrelerinin yüzeyine bağlayan ve CDC, ADCC, ADCP (Antikor bağımlı hücresel fagositoz) gibiimmün efektör mekanizmalarıyla seçici olarak fonksiyon gösteren mAb’lardır. (Lenfoma tedavisinde kullanılan anti-CD20: Rituksimab) (31) Şekil 2.9: CD20 monoklonal antikorlarının etki mekanizmaları (40) Şekil 2.9’da: CD20 mAb’ları çeşitli şekillerde tümör ölümlerine sebep olabilirler. 1: Apoptozis/ ligand blokajı: Klasik olmayan apoptozisin indüksiyonu aracılığıyla hücre ölümü ile sonuçlanan CD20 mAb’larının direkt bağlanması çoklu CD20 moleküllerinin çaprazlanması ile sonuçlanır. 2: CDC: Kompleman aktivasyonu sonucu kompleman bağımlı sitotoksisite 3: ADCC: İmmün efektör hücrelerde eksprese edilen FcγR reseptörleri tarafından opsonize tümör hücrelerinin tanınması antikor-bağımlı hücrearacılı sitotoksisiteyi başlatır. 4: Gelişmiş kompleman ADCC: FcγR yalnızca çapraz platform olarak hizmet eder ve böylece tümör hücrelerinde antijen sinyalizasyonunu artırabilir. 5: Crosslinking: Başlatılan antikor-kompleman aktivasyonu gelişmiş 32 kompleman-ADCC denilen bir süreçte kompleman reseptörleri tarafından tanımlama yoluyla tümör ölümlerini artırabilen bölünmüş kompleman parçalarını verir (40). Dördüncüsü, üç fonksiyonlu (Bispesifik) mAb’lardır. İki farklı antijen bağlarken diğer fonksiyon immün efektör mekanizmalarını çekme yeteneğindedir. (Katumaksomab, bir anti-CD3’tür. EpCAM (+), epitelyal hücre adhezyon molekülü, neoplazmlı hastalarda kötü huylu assit toplanmaları tedavisinde kullanılır.) (31) Şekil 2.10. Katumaksomab’ın öne sürülen etki mekanizması (41) Şekil 2.10’da: Üç işlevli antikor katumaksomab çeşitli immün hücreleri tarafından tümör hücrelerinin tanınmasını ve yıkımını hızlandırır. ADCC, (antibody-dependent cellular toxicity) antikor bağımlı hücresel sitotoksisite; DC-CK1, (dendritic cell cytokine 1) dendritik hücre sitokini 1; IL, interlökin; IFNγ, interferon gama; TNFα, tümör nekrozis faktör alfa; LFA, lenfosit fonksiyon antijeni; NK (natural killer) doğal öldürücü hücreler; GM-CSF, (granulocyte monocyte colony stimulating factor) granülosit monosit stimüle edici faktör (41). Beşincisi, immünokonjugatlardır. (Y-ibritumomab tiuksetan ve I-tositumomab, antiCD20 mAb radyonüklit çiftleri, lenfoma tedavisinde kullanılmaktadırlar.) (31) Altıncısı, immün uyarıcı mAb’lar, kanser hücresi/immün sistem parazitlerini ve bu parazitlerin ortaya çıkardığı sinyal yolaklarını hedefleyerek tümöre özel immün cevabın gelişimine olanak sağlarlar (31). 33 Terapötik mAb’lar Avrupa ve Amerika’da hem de diğer ülkelerde pazarlama için onaylanmışlardır. Avrupa ve Amerika’da 34 tane onaylı terapötik mAb bulunmaktadır (42). Bunlardan 14 tanesi kanser için onaylıdır. Mart 2012 olarak 28 tanesi (absiksimab, rituksimab, basiliksimab, palivizumab, infliksimab, trastuzumab, alemtuzumab, adalimumab, tositumomab-I131, setuksimab, ibritumomab panitimumab, tiuksetan, omalizumab, ekulizumab, bevasizumab, sertolizumab pegol, natalizumab, golimumab, ranibizumab, kanakinumab, katumaksomab, ustekinumab, tosilizumab, ofatumumab, denosumab, belimumab, raksibakumab, ipilimumab, brentuksimab vedotin, pertuzumab) Avrupa ve Amerika’da pazarlanmaktadır. 6 tanesinin (muromonab-CD3, nebakumab, edrekolomab, daklizumab, gemtuzumab ozogamisin, efalizumab) pazarlaması durdurulmuş veya iptal edilmiştir (42). 34 Tablo 2.2: Günümüzde kanser tedavisi için onaylı monoklonal antikorlar* (43) mAb Hedef Onaylandığı yıl Alemtuzumab (MabCampath, Campath-1H®) CD52 2001 Bevasizumab (Avastin®) VEGF 2004 Brentuksimab vedotin (Adcentis®) CD30 2011 Katumaksomab (Removab®) CD3 ve 2009 EpCAM Setuksimab (Erbitux®) EGFR 2004 Denosumab (Prolia®) RANKL 2011 Gemtuzumab ozogamisin (Mylotarg®) CD33 2000 İbritumomab tiuksetan (Zevalin®) CD20 2002 İpilimumab (Yervoy®) CTLA-4 2011 Ofatumumab (Arzerra®) CD20 2009 Panitumumab (Vectibix®) EGFR 2006 Rituksimab (MabThera®, Rituxan®) CD20 1997 Tositumomab (Bexxar®) CD20 2003 Trastuzumab (Herceptin®) HER2 1998 CTLA-4, sitotoksik T lenfosit antijen 4; EGFR, epidermal büyüme faktörü reseptörü; EpCAM, epitelyal hücre adhezyon molekülü; HER2, insan büyüme faktörü reseptörü 2: RANKL, reseptör aktifleştirici nükleus kappa ligandı; VEGF, vasküler endotelyal büyüme faktörü. * FDA veya Avrupa İlaç Ajansı tarafından bildiri Alemtuzumab kronik lenfositik lösemide; bevasizumab glioblastoma multiformede, kolorektal, böbrek ve akciğer kanserinde; brentuksimab vedotin Hodgkin ve anaplastik büyük hücreli lenfomada (Monometil auristin e ile birlikte); katumaksomab EpCAM pozitif malign assitli hastalarda; setuksimab kolorektal ve baş-boyun kanserinde; denosumab, meme ve prostat kanserinde; ibritumomab tiuksetan non-Hodgkin lenfomada ( 90 Y veya 111 In ile birlikte); ipilimumab melanomada; ofatumumab kronik lenfositik lösemide; panitumumab kolorektal kanserde; rituksimab non-Hodgkin lenfomada ve kronik lenfositik lösemide; tositumomab non-Hodgkin lenfomada (tek başına veya 131 I ile birlikte); trastuzumab meme kanserinde ve mide veya gastroözofageal adenokarsinomada endikedir (43). 35 Tablo 2.3. Faz III-IV çalışmalarda kanser tedavisi için değerlendirme aşamasında olan monoklonal antikorlar* (31) mAb Hedef Endikasyon(lar) Ch14.18 GD2 -Nöroblastoma Ganitumab IGF1R -Pankreasın metastatik adenokarsinoması Nesitumumab EGFR -NSCLC EGFR -Nazofaringeal kanser -HNC -Bölgesel ileri özofagus kanseri -Özofagus yassı epitel hücreli karsinom Ramusirumab VEGFR2 -Metastatik mide veya gastroözofageal adenokarsinoma -1.basamak sorafenib tedavisi sonrası hepatoselüler karsinoma Siltuksimab IL-6 -Relaps veya refrakter multipl miyelom EGFR, epidermal büyüme faktörü reseptörü; HNC, baş ve boyun kanseri; IGFR1, insülin benzeri büyüme faktörü 1 reseptörü; IL, interlökin; NSCLC, küçük hücreli olmayan akciğer kanseri; RANKL, RANK ligand; VEGFR2, vasküler endotelyal büyüme fakötü reseptörü 2. *Ocak 2008’den sonra başlayan, bildirisi tamamlanmayan veya sonlandırılmayan. 36 3. TARTIŞMA VE SONUÇ Monoklonal antikorlarla ilgili faz çalışmaları ve faz çalışmalarından sonra onay alan monoklonal antikorlar için ise etkinlik, güvenilirlik çalışmaları yıllardır yapılmaktadır. Bugüne kadar yüzlerce çalışma yapılmıştır ve yapılmaya devam etmektedir. Burada birkaç çalışma örnek olarak gösterilmiştir. Osteoborg A. ve arkadaşları 1996’da yayımlanan çalışmalarında önceden kronik lenfositik lösemi için tedavi edilmemiş hastalarda Campath-1H’ın (alemtuzumab) etkinliğini araştırmayı amaçlamışlardır. Humanize CD52 mAb olan alemtuzumabı ilerleyici kronik lenfositik lösemili 9 hastada ilk basamak tedavi olarak kullanmışlar ve 5 kişiye intravenöz 4 kişiye subkütan enjeksiyonu (30mg/enj’a kadar) 18 haftalık bir sürede haftada 3 kez uygulamışlardır. 3 hastada tam gerileme ve 5 hastada kısmi gerileme gözlemlenmiştir. (Yanıt oranı %89). Tüm hastalarda kandan ve 7 hastada kemik iliğinden kronik lenfositik lösemi hücreleri temizlenmiş ve yan etkiler CMV pnömonisi gelişen bir hasta hariç hafif olarak belirtilmiştir. Tüm hastalarda lenfositopeni (B ve T hücreleri) gelişmiş fakat hematolojik toksisite çok az gelişmiştir. Yapılan bu çalışma ile Osteoborg ve arkadaşları Campath-1H’ı önceden kronik lenfositik lösemi için tedavi edilmemiş hastalarda son derece etkili ve iyi tolere edilen bir ilaç olarak değerlendirmişlerdir (44). Emile G. ve arkadaşları 2013’de yayımlanan çalışmalarında tekrarlayan yumurtalık kanserinde tek ajan olarak bevasizumabı kullanarak etkinlik ve güvenilirliğini rapor etmeyi amaçlamışlardır. Bevasizumab verilmesi için hastalarda kemoterapinin en az bir önceki aşamasından sonra hastalığın nüks etmiş olması ve hastaların gastrointestinal perforasyon ile ilişkili bir durumu olmaması şartını koşmuşlardır. Daha önceden tedavi edilen 37 hasta (platin dirençli hastalığı olan hasta sayısı 33) bu geriye dönük analize dahil edilmiş ve dozları haftada kilogram başına 2.5 mg olarak uygulamışlardır. En sık görülen yan etki %62 ile hipertansiyon olmuş, intestinal perforasyon rapor edilmemiştir. 37 Yapılan bu çalışma ile Emile G. ve arkadaşları tek ajan bevasizumabı uygun terapötik indeks ile önceden tedavi görmüş ve kemoterapinin hipertansiyon, intestinal perforasyon gibi yan etkilerini istemeyen yumurtalık kanseri olan hastalar için makul bir seçenek olarak değerlendirmişlerdir (45). Ismael G. ve arkadaşları 2012’de yayımlanan çalışmalarında transtuzumabın HER2 (+) (human epidermal growth factor receptor 2=insan büyüme faktörü reseptörü 2) erken dönem meme kanserli hastalarda subkütan ve intravenöz formülasyonlarının farmakokinetik profillerini, etkinlik ve güvenilirliğini karşılaştırmayı amaçlamışlardır. HER2 (+) bölgesel olarak ilerlemiş veya inflamatuvar meme kanserli hastalar, eş zamanlı uygulanacak olan neoadjuvan kemoterapisi (ameliyat öncesi kitle küçültme kemoterapisi) için rastgele ayrılmışlar ve trastuzumab 3 haftada bir intravenöz (8 mg/kg yükleme dozu, 6 mg/kg idame doz) ya da 3 haftada bir subkütan (600 mg sabit doz) olarak uygulanmıştır. Kemoterapi dosetakselin, fluorourasilin, epuribisinin ve siklofosfamidin 4 kürü olarak uygulanmış ve ameliyattan sonra 1 yıllık tedaviyi tamamlamak için trastuzumaba devam edilmiştir. Yapılan çalışmanın sonuçlarının değerlendirilmesiyle 5 dakika boyunca uygulanan subkütan trastuzumabın standart intravenöz uygulamasıyla aynı farmakokinetik profile, aynı etkinlik ve güvenilirliğe sahip olduğu görülmüştür. Bu nedenle Ismael G. ve arkadaşları trastuzumabı geçerli bir tedavi seçeneği olarak değerlendirmişlerdir (46). Ströhlein MA. ve arkadaşları 2011’de yayımlanan çalışmalarında peritoneal karsinomatozisli hastalarda üç fonksiyonlu katumaksomabın maksimum tolere edilen dozunu ve tolere edilebilirliğini incelemeyi amaçlamışlardır. Bu açık uçlu faz I/II çalışmasında EpCAM (+) peritoneal karsinomatozisli hastalara 4 tane ardışık katumaksomab infüzyonu verilmiştir: 0.gün: 10 µg, 3.gün: 10 veya 20 µg, 7.gün: 30, 50 veya 100 µg, 10.gün: 50, 100 veya 200 µg. Maksimum tolere edilen doz sırasıyla 0., 3., 7. ve 10. günlerde 10, 20, 50 ve 200 µg olarak rapor edilmiş ve katumaksomabın kabul edilebilir bir güvenlik profili olduğu belirtilmiştir. Tedaviye bağlı en yaygın yan etkiler ateş, kusma karın ağrısı olarak gözlemlenmiştir. Son kontrollerde 17 hastanın 11’inde (%65) ilerleme gözlenmemiş, 1 hastadan tam yanıt, 3 hastadan da kısmi yanıt alınmıştır. Stroöhlein ve arkadaşları yaptıkları bu çalışmada intraperitoneal katumaksomabı peritoneal karsinomatozisli değerlendirmişlerdir (47). hastalar için umut verici bir seçenek olarak 38 Nakagawa M. ve arkadaşları 2012’de yayımlanan çalışmalarında düşük dereceli Bhücreli non-Hodgkin lenfomalı Japon hastalarda Zevalin ile radyoimmünoterapinin (RIT-Z) etkinliğini ve yan etkilerini değerlendirmeyi amaçlamışlardır. Non-Hodgkin lenfomalı 62 hasta çalışmaya dahil edilmiştir. Yanıt, PET/CT (positron emission tomography/computed tomography=pozitron emisyon tomografisi/bilgisayarlı tomografi) ile RIT-Z enjeksiyonundan 8-12 hafta sonra değerlendirilmiştir. Genel yanıt oranı %85 olmuş ve. 37 (%60) hastada tam gerileme, 16 (%26) hastada kısmi gerileme, 4 (%16) hastada durum stabil ve 5 (%18) hastada ilerleme görülmüştür. RIT-Z’ye cevap ile kemoterapi sıklığı ve fludarabinle önceden tedavi edilme öyküsü arasında anlamlı bir ilişki bulunmuştur. RIT-Z’nin etkinliği ile hastalığın evresi, hastanın yaşı, son kemoterapiden sonra geçen zaman ve lenfoma tipi arasında anlamlı bir farklılık bulunmamıştır. RIT-Z’de 4.derece trombositopeni, hastalığın evresi ve önceden fludarabinle tedavi edilme hikayesi anlamlı olarak ilişkili bulunmuştur. Ayrıca hematotoksisite ile lenfomanın tipi, hastanın yaşı ve kemik iliği nakli hikayesi arasında anlamlı bir ilişki bulunmamıştır. Anemi kemoterapinin sıklığı, kemik iliği nakli hikayesi ve önceden fludarabinle tedavi edilme hikayesi ile anlamlı bir şekilde ilişkili bulunmuştur. Nakagawa M. ve arkadaşları yaptıkları bu çalışmayla Zevalin ile immünoterapiye yanıt oranını yüksek ve hematolojik yan etkilerin daha az ortaya çıkmasında RIT-Z’yi başarılı bir tedavi olarak değerlendirmişlerdir (48). Scagliotti GV. ve arkadaşları 2012’de yayımlanan bir faz III çalışmasında multipl myeloma veya solid tümörlerden kemik metastazı yapmış vakaların tedavisinde zoledronik aside karşı denosumabın etkinliğini ortaya koyarak akciğer kanseri hastaların sağkalım verilerini sunmayı amaçlamışlardır. Hastaları, 4 mg intravenöz zoledronik asit veya 120 mg subkütan denosumab alacak şekilde rastgele ayırmışlar. Araştırma analizi için Kaplan-Meier tahminlerini ve oransal risk modellerini kullanarak küçük hücreli olmayan akciğer kanseri (NSCLC) ve küçük hücreli akciğer kanseri (SCLC) hastaları arasındaki genel sağkalımları ortaya koymuşlar. Herhangi bir akciğer kanseri 811 hastada zoledronik aside karşı denosumabın ortaya koyduğu sağkalım 7,7 aya karşı 8,9 ay ve NSCLC hastalarında 8,0 aya karşı 9,5 ay olarak belirtilmiştir. Skuamöz hücreli karsinom hastalarındaki sağkalım ise zoledronik asit için 6,4 ay denosumab için 8,6 ay olarak bildirilmiştir. Yan etki insidansları tedavi grupları arasında dengeli bulunmuştur. (Denosumab ile tedavi edilenlerde ortaya çıkan ciddi yan etki insidansı %66.0, 39 zoledronik asit ile tedavi edilenlerde %72.9) Çene kemiği osteonekrozis insidansı gruplar arasında benzer bulunmuştur. (Denosumab %0.7, zoledronik asit %0.8) Hipokalemi ise denosumab ile tedavi edilenlerde %8.6, zoledronik asit ile tedavi edilenlerde %3.8 olarak belirtilmiştir. Bu keşif analizinde denosumab zoledronik asit ile karşılaştırıldığında metastatik akciğer kanseri hastalarında daha fazla süre genel sağkalım ile ilişkili olarak değerlendirilmiştir (49). Horning SJ. ve arkadaşları 2005’te yayımlanan çalışmalarında rituksimab tedavisi sonrası ilerleyen indolent, foliküler büyük hücreli veya B-hücreli lenfomaya dönüşmüş hastalarda tositumomab ve iyodin-131 tositumomab (131I-tositumomab)’ın güvenilirliği ve genel yanıt oranını, tam yanıt oranını, tepki süresini, ilerlemesiz sağkalımı, genel sağkalımı belirlemeyi amaçlamışlardır. 40 hastaya (24’ü rituksimaba cevapsız, 11’i 6 aydan az bir süre cevap vermiş, 5’i 6 ay veya daha uzun süre rituksimaba cevap vermiş.) bu prospektif faz II çalışmasında tüm vücut dozimetrisi baz alınarak 131I-tositumomabın terapötik dozu (trombosit başına 0.65-0.75 Gy) uygulanmıştır. Önceden uygulanan tedavi sayısının 4 olduğu ve kemoterapiye dirençli hasta oranının %59 olduğu belirtilmiştir. Sonuçlar şöyle rapor edilmiştir: Genel yanıt oranı %65, tam yanıt oranı %38. Bu oranlar önceki rituksimab tedavisiyle ilişkilendirilmemiştir. Yaklaşık 3.3 yıllık bir takiple tüm hastalarda ortalama sağkalımın 10.4 ay, cevap verenlerde ise 24.5 ay olduğu bildirilmiştir. 7 cm veya daha küçük tümörü olan 1. veya 2. derece folikülerli hastalarda genel yanıt oranı %86, tam yanıt oranı %57 olmuştur. Bu alt grupta tahmin edilen 3 yıllık ilerlemesiz sağkalım %48 olmuştur. Hastaların %50’sinde 3. veya 4. derece geçici ilik toksisitesi görülmüştür. 131 I-tositumomab, rituksimab sonrası ilerleme gösteren CD20 (+) lenfomalı hastalarda etkili olarak değerlendirilmiştir. 7 cm veya daha küçük tümörü olan 1. veya 2. derece folikülerli hastalarda 3 yılda %48 ilerlemesiz sağkalım ile genel ve tam yanıtlar oldukça yüksek oranla elde edilmiştir (50). Larson RA. ve arkadaşları 2005’te yayımlanan çalışmalarında CD33 (+) akut myeloid lösemi için antikor hedefli bir kemoterapi olan gemtuzumab ozogamisin (Mylotarg) etkinlik ve güvenilirliğini analiz etmeyi amaçlamışlardır. İlk nüksünü yaşayan CD33 (+) akut myeloid lösemili hastalar bu faz II çalışmasına dahil edilmişlerdir. Hastalara 2 haftaya ayrılmış 2 doz 2 saatlik 9mg/m2 gemtuzumab ozogamisin infüzyonu olarak verilmiş ve hastalar iyileşme, hayatta kalma, tedaviyle ortaya çıkan yan etkiler açısından değerlendirilmişlerdir. Yaş ortalaması 61 olan 271 hasta gemtuzumab ozogamisin ile 40 tedavi edilmiş, kırmızı kan hücresi ve platelet transfüzyonu olmaksızın 71 hastada (%26) nötrofil iyileşmesi ≥1500/µL, hemoglobin ≥9 g/dL olarak belirtilmiştir. Trombosit iyileşmeli (≥100.000/µL) tam gerilem veya tam trombosit iyileşmesi olmaksızın (<100.000/µL) tam gerileme, sırasıyla 35 hastada (%13) ve 36 hastada (%13) gözlenmiştir. Yine sırası ile nüks gerçekleşmeden ortalama sağkalım 6,4 ay ve 4,5 ay olarak belirtilmiştir. 3. veya 4. derece nötropeni insidansı (%98) ve trombositopeni (%99) insidansı beklenen insidanslar olmasına rağmen 3. veya 4. derece sepsis (%17) ve pnömoni (%8) insidansları oldukça az bulunmuştur. 3. veya 4. derece hiperbilirubinemi, hepatik aspartat aminotransferaz, hepatik alanin aminotransferaz yükseklikleri sırasıyla %29, %18 ve %9 olarak rapor edilmiştir. Gemtuzumab ozogamisin tedavisi sonrası, hastaların %9’unda (tedavi öncesinde veya sonrasında hemapoietik kök hücre nakli olmayan) hepatik venooklüzif hastalık gelişmiştir. Sonuç olarak CD33 (+) akut myeloid lösemili hastaların ilk nükslerinde gemtuzumab ozogamisin uygulandığında, tek ajan gemtuzumab ile indüklenen %26 gerileme oranıı kabul edilebilir bir güvenlik profili olarak değerlendirilmiştir (51). Sonuç olarak, hibridoma teknolojisinin gelişmesiyle 1975 yılında hayatımıza giren mAb’lar gerek tanı gerekse tedavi konusunda özellikle de kanserde yeri doldurulamaz hale gelmiştir. Tümör immünolojisinin de daha iyi anlaşılmaya başlanmasıyla yaklaşık son 20 yılda geliştirilen, yapılan faz çalışmalarıyla kanser tedavisi için onay alan mAb’ların sayısı 14’e ulaşmıştır. Onaylandıktan sonra da ticari preparatları üzerinden de çok sayıda etkinlik ve güvenilirlik açısından klinik çalışmalar yapılmıştır. Sonuçlar üzerinden yapılan değerlendirmeler olumlu yönde olmuştur. Yeni geliştirilen monokloanl antikorlar için faz I/II ve faz III/IV çalışmaları son hızla devam etmektedir. 41 KAYNAKLAR 1. Çırakoğlu B. Bağışıklık, Bilim ve Teknik Dergisi, Mart Eki, 2003; (424): 2-5. 2. Akgün Y. İmmünolojiye giriş ve antijen. In: Serter N. (ed) Mikrobiyoloji, Anadolu Üniversitesi Açıköğretim Fakültesi Yayınları, 1996: 132-145. 3. Akgün Y. İmmünglobulinler (Antikorlar). In: Serter N. (ed) Mikrobiyoloji, Anadolu Üniversitesi Açıköğretim Fakültesi Yayınları, 1996: 164-181. 4. Candaş D. Monoklonal antikorlar, Bilim ve Teknik Dergisi, 2002; (410): 50-54. 5. Alhan E. Monoklonal Antikor Tekniği ile Çocukluk Çağı Akut Lenfoblastik Lösemisinin İmmünolojik Sınıflandırılması, Uzmanlık Tezi, Çukurova Üniversitesi Tıp Fakültesi Çocuk Sağlığı ve Hastalıkları Anabilim Dalı, Adana 1990: 59. 6. Alabaş A. İmmün Sistem, Bitirme Tezi, Ege Üniversitesi Diş Hekimliği Fakültesi Patoloji Birimi, İzmir 2006: 49. 7. Akgün Y. İmmün sistemin yapısı. In: Serter N. (ed) Mikrobiyoloji, Anadolu Üniversitesi Açıköğretim Fakültesi Yayınları, 1996: 147-162. 8. Beyaz F. T lenfositlerin gelişimi, fonksiyonları ve histokimyasal özellikleri. Erciyes Üniversitesi Veterinerlik Fakültesi Dergisi 2004; 1:(1):61-66. 9. Koz M. Farelerde Büyüme Döneminde Yapılan Egzersizin Doğal Öldürücü (Natural Killer) Hücre Sitotoksik Aktivitesine Etkisi, Doktora Tezi, Gazi Üniversitesi Sağlık Bilimleri Enstitüsü Fizyoloji Anabilim Dalı, Ankara 1996: 72. 42 10. Çetin A.Sıçanların Solunum, Sindirim ve Boşaltım Sistemlerindeki Makrofajların Histolojik Yapılarının Işık ve Elektron Mikroskobik Olarak İncelenmesi, Yüksek Lisans Tezi, İnönü Üniversitesi Sağlık Bilimleri Enstitüsü Embriyoloji ve Histoloji Anabilim Dalı, Malatya 2006: 54. 11. Cinel E. Kan ve kan bileşenlerinin immun sisteme etkisi. Yoğun Bakım Derneği Dergisi, 2005; 3:(2):56-64. 12. Erişim: [http://med.ege.edu.tr/Image/gozdoc/okuler_immunoloji_elif_demirkilinc. doc] Erişim tarihi 25 Aralık 2012. 13. Akçam FZ. HLA sistemi. Türkiye Klinikleri Tıp Bilim Dergisi, 2005; 25:829-834. 14. Yakut T. HLA doku uygunluk kompleksi, genetik lokalizasyonları ve fonksiyonları. Güncel Pediatri Dergisi, 2004; 2:53-57. 15. Holling TM, Schooten E, Den Elsen PJ. Function and regulation of MHC class II molecules in T-lymphocytes: Of mice and men. Hum Immunol 2004; 65:282-290. 16. Akın B. Kompleman Bileşeni C4A ve C4B Genlerindeki Rodgers ve Chido Belirleyicilerin Pcr, Rflp ve Dna Analizi Teknikleri ile İncelenmesi, Yüksek Lisans Tezi, Hacettepe Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı, Ankara 2007: 70. 17. Kılcı G. Ekstracorporeal Dolaşımda Priming Solüsyonunda Hydroxyethyl Starch%6 130/0.4 (voluen), Hes%10 200/0.5 ve Ringer Kullanımının Enflamatuar Yanıt ve Morbidite Üzerine Etkileri, Uzmanlık Tezi, Dokuz Eylül Üniversitesi Tıp Fakültesi Kalp ve Damar Cerrahisi Anabilim Dalı, İzmir 2006: 44. 18. Yeşilada E.Makrofaj ve lenfosit kaynaklı sitokin çalışmalarının halk ilaçlarının biyolojik etki değerlendirmesinde rolü, 14. Bitkisel İlaç Hammaddeleri Toplantısı, Bildiriler, ss 49-52, 29-31 Mayıs 2002, Anadolu Üniversitesi, Eskişehir. 19. Güner İ, Özmen D, Bayındır O. Sitokinler. Türkiye Klinikleri Tıp Bilim Dergisi, 1997; 17:65-74. 43 20. Beyaz F. B lenfositlerin gelişimi, fonksiyonları ve histokimyasal özellikleri. Erciyes Üniversitesi Veterinerlik Fakültesi Dergisi 2004; 1:(1):67-72. 21. Tuncel G. Yenidoğanda Fototerapinin IL-6 ve IL-8 Düzeyine Etkisi, Uzmanlık Tezi, Sağlık Bakanlığı Zeynep Kamil Kadın ve Çocuk Hastalıkları Eğitim ve Araştırma Hastanesi, İstanbul 2004: 59. 22. Aydın G. Deneysel Omurilik Yaralanmasında İnterlökin-10’un İnterlökin 1-Beta ve İnterlökin-6 Üzerine Etkilerinin İncelenmesi, Uzmanlık Tezi, Süleyman Demirel Üniversitesi Tıp Fakültesi Beyin Cerrahi Anabilim Dalı, Isparta 2007: 53. 23. Örnek O. IL-12 ve IL-15 ile Uyarılmış İnsan NK Hücrelerinin Kolon Karsinomuna Yönelik Anti-Tümoral Etkileri, Doktora Tezi, İstanbul Üniversitesi Sağlık Bilimleri Enstitüsü İmmünoloji Anabilim Dalı, İstanbul 2007: 55. 24. Yılmaz N. Deneysel Kolit Modelinde Anti GM-CSF’nin Etkileri, Uzmanlık Tezi, Erciyes Üniversitesi Tıp Fakültesi Genel Cerrahi Anabilim Dalı, Kayseri 2006: 69. 25. Kırıkçı AD. Kronik Hepatit B Hastalarında İnterferon Alfa-2b Lamivudin Kombinasyon Tedavisinin Etkinliği, Uzmanlık Tezi, Yüzüncü Yıl Üniversitesi Tıp Fakültesi Enfeksiyon Hastalıkları ve Klinik Mikrobiyoloji Anabilim Dalı, Van 2008: 57. 26. Erkan F. Tümör immünolojisi. Endoskopi Dergisi, 1997; (4):54-56. 27. Aslan G. Tümör immünolojisi. Türk İmmünoloji Dergisi, 2010; 15:7-13. 28. Weiner LM, Surana R, Wang S. Antibodies and cancer therapy: versatile platforms for cancer immunotherapy. Nat Rew Immunol 2010; 10(5):317-327. 29. Erişim: [http://faculty.madisoncollege.edu/mljensen/BloodBank/lectures/blood_ bank_antigens_and_antibodi.htm] Erişim tarihi 16 Şubat 2013. 30. İzgür M, Diker KS. Monoklonal antikorlar. Ankara Üniversitesi Veterinerlik Fakültesi Dergisi 1984; 31(1):98-106. 31. Galluzzi L, Vacchelli E, Fridman WH, et al. Monoclonal antibodies in cancer therapy. OncoImmunology 2012; 1(1):28-37. 44 32. Koçoğlu T, Yalçınkaya T. Rekombinant DNA teknolojisi. Mikrobiyoloji Bülteni 1992; 26:177-188. 33. Çırakoğlu B. Monoklonal antikorlar, Bilim ve Teknik Dergisi, Mayıs Eki, 2002; (414): 6-7. 34. Bulut Öner F. Biyoteknolojinin tıpta ve eczacılıktaki uygulamaları, Meslek İçi Sürekli Eğitim Dergisi, 2002; (3,4): 42-53. 35. Tuncer M. Affinite kromatografisi. In: Protein Saflaştırma-1: Kromatografik Teknikler, Cem Matbaacılık, Mersin, 2008: 159-197. 36. Aslankaraoğlu Akçael E. Poliester Fiber Destekli Dolgulu Reaktörlerde Hibridoma Kültürü ve Monoklonal Antikor Üretimi, Doktora Tezi, Hacettepe Üniversitesi Fen Bilimleri Enstitüsü Kimya Mühendisliği Anabilim Dalı, Ankara 2006: 201. 37. Erişim: [http://www.ncbi.nlm.nih.gov/pubmed/23507041] Erişim tarihi 19 Mart 2013 38. Dubois EA, Cohen AF. New drug mechanisms. Br J Clin Pharmacol 2009; 68:(4):482-483. 39. Alvarez RH, Guarneri V, Icli F, et al. Bevacizumab treatment for advanced breast cancer. The Oncologist 2011; 16:1684-1697. 40. Boross P, Leusen JHW. Mechanisms of action of CD20 antibodies. Am J Cancer Res 2012; 2:(6):676-690. 41. Linke R, Klein A, Seimetz D. Catumaxomab clinical development and future directions. Landes Bioscience 2010; 2:(2):129-136. 42. Reichert JM. Marketed therapeutic antibodies compendium. Landes bioscience 2012; 4:(3):413-415. 43. Vacchelli E, Eggermont A, Galon J, et al. Monoclonal antibodies in cancer therapy. OncoImmunology 2013; 2:(1):e22789. 45 44. Osteoborg A, Fassas AS, Anagnostopoulos A, et al. Humanized CD52 monoclonal antibody Campath-1H as first-line treatment in chronic lymphocytic leukaemia. Br J Haematol 1996; 93:(1):151-153. 45. Erişim: [http://www.ncbi.nlm.nih.gov/pubmed/23474345] Erişim tarihi 21 Mart 2013 46. Ismael G, Hegg R, Muehlbauer S, et al. Subcutaneous versus intravenous administration of (neo)adjuvant trastuzumab in patients with HER2-positive, clinical stage I-III breast cancer (HannaH study): a phase 3, open-label, multicentre, randomised trial. Lancet Oncol 2012; 13:(9):869-878. 47. Ströhlein MA, Lordick F, Rüttinger D, et al. Immunotherapy of peritoneal carcinomatosis with the antibody catumaxomab in colon, gastric, or pancreatic cancer: an open-label, multicenter, phase I/II trial. Onkologie 2011; 34:(3):101-108. 48. Nakagawa M, Uike N, Choi I, et al. Efficacy and safety of yttrium-90 ibritumomab tiuxetan in Japanese patients with non-Hodgkin lymphoma. Japanese Journal of Radiology 2012; 30:(8):642-647. 49. Scagliotti GV, Hirsh V, Siena S, et al. Overall survival improvement in patients with lung cancer and bone metastased treated with denosumab versus zoledronic acid: subgroup analysis from a randomized phase 3 study. Journal of Thoracic Oncology 2012; 7:(12):1823-1829. 50. Horning SJ, Younes A, Jain V, et al. Efficacy and safety of tositumomab and iodine-131 tositumomab (Bexxar) in B-cell lymphoma, progressive after rituximab. Journal of Clinical Oncology 2005; 23:(4):712-719. 51. Larson RA, Sievers EL, Stadtmauer EA, et al. Final report of the efficacy and safety of gemtuzumab ozogamicin (Mylotarg) in patients with CD-33 positive acute myeloid leukemia in first recurrence. Cancer 2005; 104:(7):1442-1452. 46 ÖZGEÇMİŞ KİŞİSEL BİLGİLER Adı, Soyadı: Özge ATEŞLİ Uyruğu: Türkiye Cumhuriyeti (TC) Doğum Tarihi ve Yeri: 18 Temmuz 1990, Erzincan Medeni Durumu: Bekar Tel: 0 506 947 75 52 E-mail: ozgetsl@hotmail.com EĞİTİM Derece Kurum Mezuniyet Tarihi Lise Bafra Kızılırmak Anadolu Öğretmen Lisesi, Samsun 2008 Erciyes Üniversitesi Eczacılık Fakültesi, Kayseri 2013