Beyin görsel bilgiyi 3 aşamada analiz eder
Düşük-seviye lokal kontrast, oryantasyon, renk ve hareket
gibi görsel özelliklerin ayırt edilmesi
Ara-seviye görsel sahnenin planı, yüzey özellikleri,
önplanın arkaplandan ayırt edilmesi
Yüksek-seviye nesne tanıma
Retinada Görsel İşlenme
Işık kornea ve lens tarafından odaklanır, vitreus humorden geçerek
fotoreseptörlere ulaşır. Pigment epitelindeki hücrelerde bulunan melanin
retinanın yakalayamadığı ışığı absorbe eder. Işığın gözün arkasından retinaya
tekrar yansıması sonucu görüntünün bozulması önlenir. Fotopigmentlerin
yenilenmesinde ve fotoreseptör disklerinin fagositozunda önemli rol oynar.
Foveada proksimal retinal nöronların aksonları kenara kaymış olduğu için
fotoreseptörlere ulaşan görüntüdeki deformasyon en azdır.
İki tip fotoreseptör vardır: Koniler ve basiller
Koniler basillerin 20 de biri olmasına rağmen uzaysal çözünürlükleri iki
nedenle daha iyidir: 1. Konverjansın az olması 2. Görüntünün en az
bozulmaya uğradığı foveada bulunmaları.
En düşük ışık seviyesinde yalnız basiller aktiftir = skotopik görme
Koniler “starlight” ışık seviyesinde görsel algıya katılmaya başlarlar.
Aydınlık arttıkça konilerin katkısı artar. Oda ve güneş ışığında yalnız koniler
görsel algıyı oluşturur = fotopik görme
Alacakaranlıkta hem basil hem de koniler aktiftir = mezopik görme
Dış segment fototransdüksiyon için özelleşmiştir. Işığı absorbe eden görme
pigmentlerini içerir. Dış segmentte membranın katlanması ile oluşan diskler
bulunur. Dış segment devamlı yenilenir, yeni diskler oluşurken eski diskler
pigment epitel hücrelerinin fagositik aktiviteleri aracılığı ile uzaklaştırılır.
İç segment hücrenin çekirdeğini ve biyosentetik yapılarını içerir.
Sinaptik terminal hedef hücrelerle bağlantı kurar.
Fototransdüksiyon
Karanlıkta cGMP konsantrasyonu yüksektir.
cGMP-kapılı kanallardan sodyum iyonlarının hücreye
girmesi ile hücre depolarize durumda tutulur.
Fototransdüksiyon üç evrede oluşur:
(1) Işık görme pigmentlerini aktifler; (2) bu aktiflenmiş
moleküller cGMP fosfodiesterazı uyarır; (3) cGMP
konsantrasyonundaki azalma sonucu cGMP-kapılı
kanalların kapanması ile fotoreseptör hiperpolarize olur.
Evre 1: Işık pigment moleküllerini aktifler
g
Basillerdeki görme pigmenti rodopsindir.
Rodopsin =
Opsin +
Retinal (A Vit. derivatı)
(protein)
(ışığı-absorbe eden kısım)
11-sis retinal
ışık
all-trans retinal
opsin
metarodopsin II
Metarodopsin II opsin + all-trans retinal
All-trans retinal pigment epitel hücrelerine taşınır ve all-trans
retinole (A Vitamini, 11-sis retinalin prekürsörü)
dönüştürülerek basillere geri taşınır.
Konilerde görme pigmenti = Koni opsini + 11-sis retinal
Her bir koni pigmenti koni opsininin farklı bir izoformunu
içerir.
Evre 2: Pigment moleküllerinin aktiflenmesi cGMP
konsantrasyonunu azaltır
Işık ile pigment moleküllerinin aktiflenmesi bir G
proteinini uyararak (basillerdeki transdusindir) cGMP
fosfodiesterazı aktifler. Bu enzim cGMP’yi 5’GMP’ye
parçalar.
İnaktif transdusine GDP bağlıdır. Aktif rodopsin ile
etkileşince GDP’yi GTP ile değiştirir ve aktiflenir.
Tek bir rodopsin molekülünün bir foton ile aktivasyonu
saniyede 105 cGMP molekülünün yıkılmasına neden olur
yüksek sinyal amplifikasyonu
Bir rodopsin 102 trandusini aktifler,
Her bir transdusin Bir fosfodiesterazı aktifler,
Bir cGMP fosfodiesteraz saniyede 103 cGMP’yi hidrolize eder.
Işığa yanıt iki mekanizma ile sonlanır:
1. Trandusin GTP’yi hidrolize ederek (GTPaz aktivitesi
ile) kendisini inaktif duruma getirir.
2. Aktiflenmiş rodopsin opsin kinaz ile fosforillenir.
Fosforillenmiş rodopsin düzenleyici bir protein olan
arrestin ile etkileşerek inaktiflenir.
Evre 3: cGMP konsantrasyonunda azalma cGMP-kapılı
iyon kanallarını kapatır, fotoreseptör hiperpolarize olur
Karanlıkta iki akım baskındır: cGMP
kapılı kanallardan içeri doğru & kapısız
K+ kanallarından dışarı doğru akım.
Na+-K+
pompası
Na+
ve
K+
konsantrasyonlarını belirli seviyede tutar.
Karanlıkta membran potansiyeli -40 mV
civarıdır.
Işıkta cGMP kapılı kanallardan içeri
doğru olan akım azalır ve hücre
hiperpolarize olur.
Işığın şiddetine göre hiperpolarizasyon
-40 ila -70 mV arasıdır.
Fotoreseptörler ışık şiddetindeki değişikliklere yavaş
adapte olurlar
Karanlığa veya aydınlığa adaptasyon sırasında konilerde iki
önemli değişiklik oluşur: Işığa adaptasyon
1. Membran potansiyelinin yavaşça eski haline dönmesi: Parlak bir
ışık tüm cGMP kapılı kanalları kapatır (-70mV) eğer ışık
devam ederse membran -70 ila -40 mV arası bir potansiyele
depolarize olur daha yüksek şiddette bir ışığa yanıt verebilir.
2. Reseptörün duyarsızlaşması: Uzun süren bir ışıkta, membran
potansiyelinde değişiklik oluşturabilen ışık şiddetindeki fark artar.
Konilerin dış segmentinde Ca+2 un azalmasına bağlı
Kalsiyum fototransdüksiyon
işlevini düzenler;
yolağında
çeşitli
proteinlerin
Karanlıkta Ca+2 cGMP-kapılı kanallardan dış segmente girer; bir
taşıyıcı ile dışarı atılarak konsantrasyonu belirli seviyede tutulur.
Uzun süren bir aydınlanmada; cGMP-kapılı kanallar kapanır Ca+2
girişi azalır çıkış devam ettiğinden Ca+2 konsantrasyonu azalır.
1. Ca+2 un guanil siklaz üzerindeki inhibe edici etkisi azalır
cGMP sentezi yavaşça artar cGMP-kapılı kanallar yeniden açılır
koniler depolarize olur.
2. a) Görme pigmentlerinin inaktivasyonu hızlanır, böylece bir ışık
uyaranının cGMP fosfodiesterazı aktive edici etkisi azalır.
b) cGMP-kapılı kanalların cGMP değişikliklerine duyarlılığı azalır.
Aynı sayıda cGMP-kapılı kanalın kapanabilmesi için daha şiddetli
bir ışık gerekir
Retinanın çıktısı gangliyon hücreleri tarafından taşınır
Fotoreseptörler ışığa membran potansiyellerindeki kademeli
değişiklik ile yanıt verirler. Gangliyon hücreleri ise ışık
bilgisini aksiyon potansiyelleri ile taşırlar.
Fotoreseptörler ile gangliyon hücreleri arasında üç tip
aranöron vardır: bipolar, horizontal ve amakrin hücreler.
Fotoreseptörlerden gelen sinyalleri birleştirirler.
Basil ve koni sinyalleri aynı gangliyon hücresinde sonlanır.
Herbir gangliyon hücresi aydınlanmanın seviyesine göre hem
koni hem de basil girdilerine yanıt verir.
Fotoreseptörler dış nükleer, aranöronlar iç nükleer, gangliyon
hücreleri ise gangliyon hücre tabakasında bulunurlar.
Fotoreseptörler, bipolar ve horizontal hücreler dış pleksiform
tabakada; bipolar, amakrin ve gangliyon hücreleri ise iç pleksiform
tabakada sinaps yaparlar.
Gangliyon hücresinin reseptif alanı
Gangliyon hücresinin monitörize ettiği retina alanıdır.
--- Kabaca daireseldir.
--- İki bölüme ayrılır: merkez (center) ve çevre (surround).
Gangliyon hücrelerinin reseptif alanı foveada küçük, retina
periferinde geniştir.
Gangliyon hücreleri reseptif alanlarının merkez ve
çevresinin farklı aydınlanmasına optimal olarak yanıt verir.
Reseptif alanlarının merkezine uygulanan ışığa verdikleri
yanıta göre iki tip gangliyon hücresi vardır: On-center ve
off-center hücreler
Her iki tip hücrede de
reseptif alanın çevresine
uygulanan ışık, merkeze
uygulanan ışığın
oluşturduğu yanıtı ortadan
kaldırır. Bu nedenle tüm
reseptif alanın difüz
aydınlanması yalnızca
küçük bir yanıt oluşturur.
On-center ve off-center
hücreler kabaca eşit
sayıdadır. Her fotoreseptör
ikisine de output gönderir.
Görsel bilginin işlenmesi
için iki paralel yolak oluşur.
Merkez-çevre organizasyonu görme alanındaki kontrast
bilgisinin tespit edilmesi için bir adaptasyondur.
Nesnelerin algılanması ışık kaynağının şiddetine değil
nesne ve arka planı arasındaki kontrasta dayanır.
Bir gangliyon hücresinin ateşleme oranı merkez ve
çevreyi aydınlatan ışığın şiddetleri arasındaki farkın bir
ölçümüdür.
Paralel on-center ve off-center yolaklar görsel sistemin
performansını arttırır çünkü her bir gangliyon hücresi
aydınlanmadaki ya hızlı artışa ya da azalmaya en iyi yanıt verir.
On-center hücrenin ateşleme oranı alaca karanlıkta düşüktür;
ateşlemedeki hızlı artış, merkezde ışık şiddetinin hızla arttığını
gösterir.
Off-center hücrenin ateşleme oranı ışıkta düşüktür; ateşlemedeki
hızlı artış, merkezde ışık şiddetinin hızla azaldığını gösterir.
Gangliyon hücreleri iki sınıftır: magno ve parvo. Her bir sınıf
hem on-center hem de off-center hücreleri içerir.
Aydınlanma uzun sürerse on-center gangliyon hücresinin deşarjı
azalır. Aydınlanma azalırsa zaten bu hücrelerin düşük olan
deşarjının daha da azalması güvenilir bir sinyal olmaz. Halbuki
off-center hücreler bu uyarıya ateşleyerek yanıt verirler.
On- ve off-center bipolar hücreler tek bir koniden gelen sinyal için
paralel yolaklar oluşturur
Her bir koni hem on-center hem de offcenter bipolar hücrelerle sinaps yapar. Her
bir bipolar hücre aynı tip gangliyon
hücresi ile sinaps yapar.
Karanlıkta koniler depolarize olduğundan
glutamat salınır. Glutamat on-center
bipolar
hücreleri
inhibe
eder
(hiperpolarizasyon, K+ kanallarını açar
veya Na+ kanallarını kapatır), off-center
bipolar hücreleri uyarır (depolarizasyon,
Na+ kanallarını açar).
Işıkla koniler hiperpolarize olunca
glutamat salınımı azalır. On-center
hücreler depolarize, off-center hücreler
hiperpolarize olur.
Off-center bipolar hücrelerde glutamatın
iyonotropik reseptörleri (AMPA ve kainat)
var. Fotoreseptörden salınan glutamat
depolarizasyona neden olur signconserving
On-center bipolar hücrelerde G-proteinle
eşleşen metabotropik glutamat reseptörleri
(mGluR6) var. Glutamat ile bağlanınca
cGMP-kapılı Na+ kanallarını kapatan bir
kaskadı aktifler. Hiperpolarizasyon olur
sign-inverting
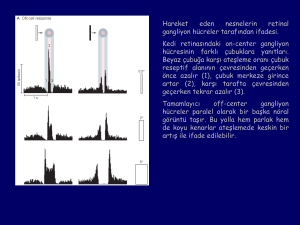
On-center gangliyon hücresinin spot ışığına karşı deşarj oranıspotun reseptif alanının merkezinden uzaklığının bir
fonksiyonu olarak. X eksenindeki sıfır merkeze karşılık gelir.
Reseptif alanları aydınlık-karanlık bir kenar boyunca dağılan
hipotetik bir grup on-center gangliyon hücresinin yanıtları.
Aktivitesi en fazla etkilenen nöronlar reseptif alanları
aydınlık-karanlık sınırı boyunca uzananlardır (B ve D).
Tek bir on-center gangliyon hücresinin reseptif alanının merkezindeki küçük bir ışık
noktasına karşı deşarj oranı. Her bir eğri sabit bir arkaplan aydınlanma seviyesinde (en
yükseği 0) farklı şiddetteki noktalar ile uyarılan deşarj oranını gösteriyor. Yanıt oranı 1
log birim aralığı boyunca uyaran şiddetine orantılıdır ama işlediği aralık arkaplan
aydınlanması arttıkça sağa kayar.
Konilerden gangliyon hücrelerine bilgiler iki yolakla
iletilir
Bipolar hücreler konilerden gelen sinyalleri gangliyon
hücrelerine ya doğrudan (reseptif alanın merkezindeki koniler)
ya da özellikle horizontal hücreler aracılığı ile dolaylı olarak
(reseptif alanın çevresindeki koniler) taşırlar.
Horizontal hücreler uzaktaki konilerden gelen bilgileri, bipolar
hücreye, bipolar hücrenin reseptif alanının merkezindeki
konilere taşıyarak iletirler.
Bipolar hücrelerin reseptif
organizasyonuna sahiptir.
alanları
da
merkez-çevre
On-center bipolar hücrede
merkez-çevre antagonizması:
Horizontal hücre hem on-center
bipolar hücrenin çevresindeki bir
koniden input alıyor hem de
bipolar hücrenin reseptif alanının
merkezindeki postsinaptik bir
koni ile bağlantı yapıyor.
Bipolar hücrenin çevresindeki koniler ışık uygulanınca hiperpolarize
olur postsinaptik horizontal hücre hiperpolarize olur horizontal
hücre tarafından salınan inhibitör transmitterin salınımı azalır reseptif
alanın merkezindeki koniler depolarize olur on-center bipolar
hücrenin hiperpolarize olmasına neden olur.
Fotoreseptörden salınan
glutamat horizontal hücrede
depolarize edici (signconserving), horizontal hücreden
salınan GABA fotoreseptör
terminalinde hiperpolarize edici
(sign-inverting) etkiye sahiptir.
Horizontal hücre ağından
inputların net etkisi
fotoreseptörün membran
potansiyelinde
fototransdüksiyon ile indüklenen
değişikliklere karşıttır.