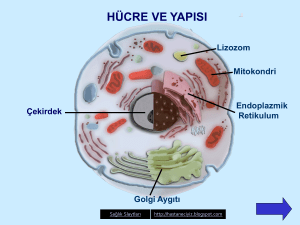
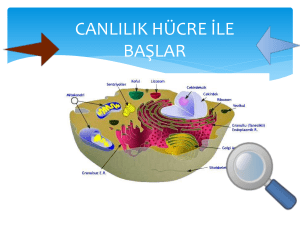
Sitopazma Organelleri Endoplazmik Retikulum Hücre bölünmeleri
advertisement

Organeller ve Protein Sentezi Hücre-içi Zar Sistemi ve Sitoplazma Prokaryot hücrelerde bulunmayan bir hücre-içi zar sisteminin ökaryot hücrelerde gelişmiş olduğu elektron mikroskobu ile gözlenmiştir. Bu zar sistemi ya paralel zarlar, keseler, vakuoller şeklinde olabilen özelleşmiş zarlı bölgelerdir veya belli organellerin etrafını bir zar örtü çevirmiştir. Buna topluca hücre-içi zar sistemi veya sitoplazmik vakuol sistemi denilmektedir. Bu zar sistemi ile hücrenin içi birbirinden bağımsız çalışabilen kompartımanlara ayrılmış bulunmaktadır. Bu zar sistemi başlıca endoplazmik retikulum, Golgi kompleksi ve çekirdek zarından oluşmaktadır. Sitoplazma Hücrenin en önemli bir kısmıda sitoplazmadır. Işık mikroskobunda az çok homojen gözüken sitoplazma gerçekte oldukça heterojen; kolloid bir sistemdir. Hücre-içi zar sisteminin dışında kalan aralıkları dolduran kısım sitoplazmayı teşkil eder. Gerçek bir iç çevre meydana getiren sitoplazmaya sitoplazmik matriks (dolgu maddesi) de denir. Hücrenin bir çok kolloid özellikleri sitoplazmik matriksle sağlanır. Örneğin amöboid hareket, iğ teşekkülü, bölünme, hücre içi hareket (sikloz), vizkozite değişiklikleri sitoplazmadaki kolloit sistemin jel-sol transformasyonları ile meydana gelir. Sikloz, kasılma gibi hareketlere hücre zar sistemi de katılır. Sitoplazmik matriks hücreye şeklini verir, hücreyi destekler ve hücre bölünmesi, hücre hareketi, sil ve kamçı hareketi gibi hücre faaliyetlerinde görevlidir. Sitoplazmanın zemin maddesi sitoplazmik flamentler ve mikrotüpcük adı verilen üç boyutlu ince bir iplik ağından oluşmuştur. Bu ağın aralıkları su ile doludur. Suyun içinde inorganik iyonlar ile küçük moleküller (örneğin; enzimler, vitaminler, diğer proteinler ve şekerler) erimiş halde bulunurlar. Ağın en önemli maddesi proteindir. Bu polimer ağın yapısında yüzden fazla protein çeşidinin bulunduğu elektroforezle gösterilmiştir. Polimer ağı kenarlarda hücre zarına bağlanır ve mitokondriler, endoplazmikretikulum, ribozomlar, polizomlar gibi birçok organeli birbirine bağlar. Ayrıca hücre iskeletini teşkil eder. Katı ve statik bir yapı değildir. Hücre faaliyetine ve şeklindeki değişikliklere göre değişir. Sitopazma Organelleri Endoplazmik Retikulum Hücre bölünmeleri sonucu meydana gelen genç hücrelerde sitoplazma hemen tamamen bağımsız haldeki ribozom ve polizomlardan ibarettir. Bu durumdaki hücreler sadece kendileri için çalışırlar. Ancak hücreler belli bir olgunluk düzeyine ulaşınca organizmanın bütününü ilgilendiren faaliyetlere de katılmak zorundadırlar. Hücrelerin çok çeşitli maddeler yapıp salgılamaları da bu amaca yöneliktir. İşte bu maddeleri sentezlemek üzere, sitoplazmada endoplazmik retikulum belirmeye başlar. Endopazmik retikulum direkt olarak hiyaloplazmadan gelişebilirse de, daha çok çekirdek dış zarından köken alır. Onun için de çekirdek zarı ile olan bağlantılarına sık sık rastlanır. Buna karşılık hücre zarı ile direkt bağlantısı çok az hücre türünde –örneğin; kas hücrelerinde- görülebilir. Olgun alyuvarlarla, trombositler ve bakterilerde endoplazmik retikulum bulunmaz. bunların dışındaki bütün hayvansal ve bitkisel ökaryot hücreler bu organale sahiptir. Endoplazmik retikulum üzerinde küçük, yoğun ribonükleoprotein partikülleri bulunur. Bunlar ribozomlardır. Endoplazmik retikulum zarları hücre zarı gibi birim zar yapısındadır. Kalınlığı ortalama 50-60 A° olup hücre zarından incedir. Hücre zarında olduğu gibi, kutupsuz yağ asidi kuyrukları birbirine dönük duran iki lipittabakası ile lipitlerin üzerinde veya arasında yer alan proteinlerden yapılmıştır. Endoplazmik retikulum, memebranları üzerinde ribozomların bulunup bulunmamasına göre isim alır. a) Granüllü Endoplazmik Retikulum (GER) b) Granülsüz (Granüler) Endoplazmik Retikulum (DER) olmak üzere iki türü vardır. Granüllü Entoplazmik Retikulum: Üzerinde taneciklerin bulunması nedeniyle GER denir. Hücrelerin çoğunluğu bu tür retikulum taşır. Bunun miktarı hücreden hücreye değişebiliceği gibi, hücrenin fonksiyon durumuna göre de artıp eksilir. GER, hücrenin protein sentezi ile ilgili organelidir. Kuvvetli protein biosentezi yapan hücrelerde (bez epitel hücreleri, fibroblastlar, sinir hücreleri vb.) daha bol olarak bulunurlar. Protein sentezinin yapım yerleri ribozomlardır. Özellikle salgı yapan hücrelerde sayıları fazla olan ribozomların bazı yerlerde zincirler ve sarmaller biçiminde toplu halde bulundukları görülür, bunlara polizom veya poliribozom denir. Ribozomların polizomlar halinde toplu bulunuşu protein sentezini kolaylaştırır. Ribozomlarda protein sentezi yapıldıktan sonra, yapılan proteinler ER’ma ait keselerin boşluklarına geçerler. Boşluklara geçen proteinler burada tekrar fabrikasyona uğrar ve daha yüksek kuruluşta olan proteinlere donüştürülürler. Bu proteinler GER kesecikleri içinde ya homojen ve az yoğun bir kitle halinde, ya da değişik çapta olan yoğun granüller halinde toplanırlar. Buralarda toplanan proteinler, kesecikler tarafından Golgi bölgesine itilirler. Golgi aygıtında bunlara daha bazı maddelerde eklenerek olgunlaşmaları sağlanır. Golgiye gelen homojen proteinler çoğu hücrelerde granül haline dönüştürülür, hücreden dışarıya verilecek olanlar bundan sonra hücre yüzeyine sevkedilirler. Granülsüz Endoplazmik Retikulum Üzerinde ribozom bulundurmayan düz endoplazmik retikulumdur (DER). Bu tür retikulum her hücrede az yada çok miktarda bulunabilir, fakat en gelişkin olduğu hücreler karaciğer eptel hücreleri, steroid hormon sagılayan iç sagı bezlerine ait epitel hücreleri, lipit sentezi yapan hücreler ve kas hücreleridir. Karaciğer epitel hücrelerinde GER ile bir arada bulunan ve yer yer onunla devam halinde bulunan DER, glikojen yapımının ara maddesi olan glikoz-6- fosfat taşır. Bu da onun glikojen yapımında rol oynıyan bir organel olduğunu gösterir. Ayrıca lipit yapımının da yine DER’ ma bağlı olduğu sanılmaktadır. Gerçektende, lipit tabiatinda olan hormonların (steroid hormonları) yapıldığı hücreler DER’ dan zengindir. Yağ hücrelerinde sadece DER bulunur. Bu tür retikulum, lipitlerin hücre içindeki transportunda da rol oynar. Endoplazmik retikulum da ister GER ister DER olsun diğer organeller gibi, sitoplazma akımına uyarak devamlı şekilde hareket eder. Hem GER hem de DER birçok hücre de beraber bulunur , fakat oranları farklıdır. ER’un görevlerini şu şekilde toparlayabiliriz; 1. 2. 3. 4. 5. Mekanik destek, Enzim etkenlikleri, Protein sentezi, Lipit sentezi, Karbonhidrat sentezi. Golgi Kompleksi Golgi kompleksi sitoplazmadaki zar sisteminin bir parçası olup başlıca zarlardan yapılmış bir organeldir. ER ile devamlılığı gösrterilmiştir. Fakat belirli bir yapı ile ER’ dan ayrılmaktadır. Üzerinde ribozom granülleri bulunmaz Elektron mikroskobu ile çift olarak uzanan yassılaşmış keselerden oluşan bir demet ile bunun çevresinde farklı büyüklükte ve çok sayıda yuvarlakça kesecikler ile büyük keselerden ve vakuollerden oluştuğu gösterilmiştir Her GK tipik olarak 3-8 adet paralel uzanan yassı keselerden yapılmıştır. Keselerin sayısı hücre tipine, göreve ve diğer faktörlere bağlıdır. İçte uzanan yassı keseler dışardakilerden daha kısa olurlar. Yassı keseler hafifçe bükülmüşlerdir. Böylece yapı konkav ve konveks yüzeyleri olan konsantrik tabakalar meydana getirmiştir. Organelin çevresinde yassı keselerden gelişen küçük kesecikler bulunur. Büyüklükleri faklı olur. Bazen kesecikler yassı keselerin ucundan itibaren dışa doğru dizilmiş küçük bir dizi teşkil edebilirler. Kesecikler ve keseler genellikle yassı keselerin çevresinde görülür. Daha büyük keseler en içteki yassı tarafta yer alırlar. Bu büyük keseler, yassı keselerin içinde salgı materyali birikmesi sonucu, yassı kesenin şismesi ile teşekkül ederler. Şişen kısım zarı ile birlikte keseden kopup ayrılır. Her ne kadar Golgi kompleksinin yapısı olduka sabit ise farklı hücre tiplerinde organelin yapısı, yeri, büyüklüğü faklıdır. Hücrenin fizyolojik durumuna göre G.K nin yapısında farklar gözlenir. Hücrenin salgı aktivesine göre G.K’ nin keselerinin ve keseciklerin sayılarında azalma ve çoğalma olmaktadır. Hücrede birden fazla G.K bulunabilir. G.K genellikle çekirdeğin yakınında yer alır. Golgi kompleksi ile endoplazmik retikulum zarları arasında granüllü endoplazmik retikulumdan Golgi Kompleksine materyal taşıyan yapılar bulunur. Bunlara geçit elemanları denir. Golgi kompleksinin sentez ve salgılama işlerinde görevli olduğu gösrerilmiştir. Lipoproteinlerin sentezlenmesinde de Golgi Kompleksinin rol oynadığı gösterilmiştir. Golgi komleksinin diğer önemli bir görevi lizozomların teşekkülünde görülür. Çok sayıda hidroliz enzimleri kapsayan lizozomlar tipik salgı kesecikleri halinde teşekkül eder. Granüllü endoplazmik retikulumda sentezlenen enzimler, aynı şekilde golgi kompleksine geçerek küçük keseciklere verilir ve küçük keseciklerin birleşmesi ile lizozomlar meydana gelir. Spermatitlerde Gogi kompleksi akrozom teşekkülü ile ilgilidir. Süt bezlerinde protein granüllerinin, lökositlerdeki lizozomal granüllerin, bazı oositlerde vitellus damlacıklarının Golgi kompleksinin keselerinde paketlenerek salındığı gösterilmiştir. Böylece hücrede granüllü ve granülsüz endoplazmik retikulumda sentezlenen çeşitli maddeler Golgi kompleksine gelir ve burada bu maddelere, gerekiyorsa başka maddeler eklenir ve sentezlenen maddeler organelin uçlarından kopup ayrılan kesecikler içinde zarla çevrili granüller halinde paketlenmiş olur. Bu zar hücre zarı ile birleşme yeteneğinde olduğundan gerektiğinde içeriğini kolayca hücre dışına boşaltabilir. Mitokondriler Mitokondriler hücre içinde yapı ile görevin çok iyi bir şekilde birlikte organize olduğu, çok iyi gelişmiş bir organeldir. Mitokondrilerin ışık mikroskobunda görülen ve tipik biçimleri olan sosis şekilleri hücreden hücreye değişirsede aynı görevi yapan hücrelerde oldukça benzerlik gösterir. Mitokondrilerin şekilleri hücredeki görev safhasına göre de değişir. Canlı hücrede faz-kontrast mikroskobu ile incelenince devamlı olarak hareketli oldukları ve bir dakikadan daha kısa bir süre içinde uzadıkları, dallandıkları, bölündükleri ve birleştikleri gözlenmiştir. Bu durum organelin, hücre zarı gibi, çok dinamik olduğunu göstermektedir. Mitekondrilerin büyüklükleri değişebilir. Çoğunluk enleri 0,5 mikron olmakla beraber uzunlukları farklı olur. En fazla 1-7 mikron uzunluğunda olabilirler. Mitokondriler genel olarak, sitoplazmanın her tarafına düzgün bir şekilde dağılmışlardır. Mitoz sonrasında iğ ipliklerinin yakınında bulunurlar ve hücrenin bölünmesi ile az çok eşit sayıda oğul hücrelere dağılırlar. Mitokondrilerin hücredeki yeri enerji ihtiyacına bağlıdır. Hücrenin enerji deposu olan mitokondriler, nerede enerjiye ihtiyaç varsa daima orada bulunurlar veya enerji gerektiren bölgelere giderler. Mesela bazı kas hücrelerinde mitokondriler, kas tellerinin çevresini halka gibi sararlar. Kas hücrelerinde çok sayıda mitokondri bulunması bu hücrelerin yüksek enerji ihtiyacının bir delilidir. Mitokondriler hücrede hem aktif hem de pasif olarak sitoplazmik akıntılarla yer değiştirirler. Mitokondrilerin hücredeki sayıları hücre tipine ve organelin fizyolojik durumuna göre değişir. Bazı oositlerde 300.000 kadar mitokonri bulunduğu saptanmıştır. Genellikle bir çok hücrede 500-1000 kadar değişen bir sayıda mitokondri bulunmaktadır. Mitokondrilerin bazı görevlerini kloraplastlarınn yüklenmesi sebebiyle yeşil bitki hücrelerinde daha az mitokondri bulunduğu görülür. Işık mikroskobu ile homojen olarak görülen mitokondriler elektron mikroskobu ile çok kompleks bir zar organizasyonu gösterirler. Dıştan mitokondriyi saran 60 A° kadar kalınlıktabir zar vardır. Bu zarın iç tarafında ikinci bir zar daha bulunur ve ikinci zar mitokonrinin iç boşluğuna uzanan ve genellikle yassı keseler şeklinde olan kompleks katmanlar meydana getirir. Bu katmanlara krista adı verilir İki zarın arasında 60-80 A° kadar olabilen bir açıklık görülür. İkinci zar da 60 A° kalınlığındadır. İki zar arasında kalan aralık bir dış kompartıman meydana getirir. İç zarın iç boşluğu ikinci bir iç kompartıman teşkil eder. Burası oldukça yoğun olan ve mitokondri matriksi denilen bir materyal ile doludur. Matrikste çok çeşitli enzimler bulunur Ayrıca Diğer organellerde bulunmayan bazı yapılarında matrikste bulunduğu gösterilmiştir. Genel olarak homojen görülen matriks materyali içindeki bu yapılar oldukça yoğun ve büyük granüller, fibriller, kristalar, mikrotupcüklerdir. Büyük granüller ribozomlardan 2-3 defa büyük olan yuvarlak yapılardır. Granüllerin bir tipi depo kalsiyum iyonlarıdır. Hücre içindeki bir çok olayların düzenlenmesinde kalsiyum iyonlarının önemi çok büyüktür. Bunların yanında, mitokondri ribozomu olarak tanımlanmış çok sayıda küçük granüllerin de bulunduğu gösterilmiştir. Mitokondrilerin hücre metabolizmasının hücre içi solunum sistemindeki görevi çok önemlidir. Bu görev organelin yapısı ile sıkı ilişkisi olup zarlar arasındaki aralıklar bu sisteme giren enzimlerin ve maddelerin yerleştiği yerlerdir. Mitokondrileri hücrenin solunum merkezi olarak düşünürsek, buradaki kullanılan yakıt maddeleri, besin olarak dışarıdan alınan protein, karbonhidrat ve yağlardır. Fakat bu maddelerin molekülleri büyük olduklarından mitokondrilere giremezler. Önce sitoplazmada bulunan enzimlerin etkisi ile daha küçük moleküllere ayrılırlar. Yani proteinler, amino asitlere, karbonhidratlar pruvik asite ve yağlar da yağ asitlerine parçalanırlar. Ancak bundan sonra, bu ürünler mitokondrilerin zarlardan geçerek matrikse girerler. Bu organeller aldıkları besin maddelerini-pruvik asit, yağ asitleri ve amina asitleri-taşıdıkları oksidasyon (solunum) enzimleri ile parçalayarak büyük bir miktarda enerji açığa çıkarırlar. Bu enerjinin sağlanmasında kullanılan başlıca yakıt glükoz dur. Glükoz oksijen varlığında veya yokluğunda, glikoliz olayı ile parçalanır ve enrji verir. Olay oksijen yokluğunda olursa, buna anaerobik glikoliz veya anaerobik solunum denir. Sentezlenen enerji miktarı düşük olur (58.000 kalori) ve laktik asit meydana gelir. Laktik asitte henüz enerji vardır. Halbuki oksijen varlığında yani aerobik glikoliz olayında glikoz karbondioksite ve suya parçalanırken fazla miktarda enerji meydana gelir. Kendileri bu enerjinin çok az bir kısmını- mitokondriyal enzimlerin yapımı için- kullanır, gerisini ise hiyaloplazmaya verir. Bu enerji verişi şöyle olur: Mitokondrilere yakılıcak besin maddeleri dışında adenozin difosfat (ADP) ve fosfat (PO4 ) molekülleri de girer.Mitokondrilerde bulunan ikinci grup enzimler- bunlara fosforilasyon enzimleri denir-, her ADP molekülüne bir fosfat molekülü bağlıyarak, bunların adenozintrifosfat (ATP) haline geçmelerini sağlarlar. Mitokondrilerde üretilen enerjinin büyük bir kısmı, böylece ATP moleküllerine bağlanmış olur. ATP mitokondrilerden hiyaloplazmaya geçerek her tarafa dağılır. Hiyaloplazmada bulunan adenozintrifosfataz (ATPase) enzimi ATP' yi, ADP ve fosfat moleküllerine ayırır ve enerji bağımsız hale gelir. Hücrelerde enerji çeşitli amaçlarla kullanılır. Hücre, şeklini ve iç düzenini enerji ile sürdürebilir. Hücrede olagelen çok karmaşık biyosentez olayları da enerji ile gerçekleşir. Hücre hareketleri ve bölünmeleride enerji harcanmasını gerektirir. Bütün bunların dışında, vücut ısısı da, enerjinin ısıya dönüşmüş şeklinden başka bir şey değildir. Birkaç ilkel organizmada (hastalık yapan bakteriler) anaerobik tip solunum meydana gelir ve bu canlılara anaerob organizmalar denir. Bunlar mecburi anaeroblardır. Birçok bakteriler, bira mayası oksijen sağlanıp sağanmamasına göre, ya aerobik veya anaerobik solunum yaparlar. Bunlar da fakültatif anaeroblardır. Yüksek bitki ve hayvan hücreleri aerobik solunum yaparlar. Aerobik solunum olayını formülle kısaca aşağıdaki gibi özetlemek mümkündür; C6H12O6+6CO2= 6CO2+6H2O+ enerji (686 000 kalori) Organizmalar enerji sağlamada glukozdan başka maddeleri de kullanırlar. İnsanlar ve diğer memelilerin çoğunluğu yağ ve proteinlerin sindirim ürünleri olan yağ asitlerini ve aminoasitlerini kullanırlar. Bakteriler çok çeşitli organik bileşikleri yakıt olarak kullanmaktadırlar. Lizozomlar Bunlar da membranlı organellerdendir. Golgi kompleksi tarafından yapılırlar. Görevleri hücre içi sindirimi sağlamaktır. Lizozomlar içinde çok sayıda enzim bulunur. Bu enzimlerin özelliği hafif asit ortamda her türlü biyolojik bileşikleri parçalamalarıdır. Yani hepsi hidrolitik enzimlerdir. Lizozomlarda bulunan enzimler bir zarla çevrili olduğu için, içinde bulunduğu hücreye zararlı olmazlar. Bu enzimler canlı organizmaların temel yapılarını yani protainleri, yağları, karbonhidratları, nükleik asitleri parçalarlar. Bu temel yapılar lizozomun içine alınarak parçalanma yapılır. Golgi komplekslerinden ayrılan vakuollerden bir kısmı, hücre dışı sindirimi gerçekleştirecek olan salgı granüllerine dönüşürken, diğer kısmıda lizozomları meydana getirirler. Bu ilk lizozomlara primer lizozomlar yada inaktif lizozomlar denir. İçerikleri homojen ve genellikle yoğundur. Aşağı yukarı bir salgı granülü iriliğindedirler. Primer lizozomlar, pinositoz ve fagositoz yoluyla hücreye dışardan alınan maddelerle, metabolizma sonucu hücrede şekillenen bir kısım maddeler yada yaşlanmış hücre organelleriyle birleşir ve heterojen kuruluşta olan sekunder lizozomlara (aktif lizozomlar) dönüşür. Lizozom enzimlerinin hücreye zarar vermesini engelleyen zar hasar görürse enzimler hücre içine yayılırlar ve bütün hücrenin sindirilmesine sebep olurlar. Birim zar yapısında olan lizozom zarının az bir hasar görmesiyle zar kendini tamir eder. Lizozomlar hücrenin içindeki bazı parçaların ortadan kaldırılmasını sağlarlar. Bu olay bazı hayvanların metaforfozunda kullanılan fizyolojik bir metoddur. Metaforfaz sırasında, lizozomların enzimleri ile çok sayıda hücre ve bunun sonucu bazı dokular tahrip ve yok edilir. Mesela kurbağa metaforfozunda, kurbağa tetarisinin kuyruğu kuyruk hücrelerindeki lizozomların içnde bulunan katopsin enzimlerinin faaliyetiyle dejenere olarak ufalanır ve yok olur. Böceklerin metaforfozunda da, aynı şekilde, larvaya ait dokular bu dokuların hücrelerindeki lizozomlarda bulunan enzimlerle parçalanarak sindirilirler. Bu olaya otoliz, histoliz gibi adlar verilir. Kısaca otoliz veya histoliz hücre veya dokuların erimesidir. Lizozomlar, krmızı kan hücreleri hariç, incelenen bütün hücrelerde bulunmuştur. En tipik ve en bol olarak beyaz kan hücrelerinde granüller halinde bulunurlar. Çapları 0,2-0,6 mikron arasında değişen yapılardır. Mikrocisimler Bu organel, ufak (0.3-1.5μ), yuvarlak veya oval granüller halindedir. Unit membranla çevrilmişlerdir. Hidrojenperoksit metabolizması ile ilgili enzimleri içerirler. Peroksidaz enzimleri taşıdıklarından mikrocisimler peroksizom diye isimlendirilirler. Mikrocisimler her hücrede bulunmazlar. Karaciğer epitel hücreleri ile birkısım böbrek kanalcıklarının duvarlarını oluşturan epitel hücrelerinde bol olarak bulunurlar. Ayrıca peroksizomlar ER ve mitokondrilerle, bitkilerde ayrıca kloroplastlarla sıkı fonksiyonel ilişkileri bulunduğu ve içerdiği enzimlerden üçünün hidrojenperoksit oluşumunda rol oynadığı, dördüncü katalaz enzimininde hücre için zararlı olan peroksitin su ve oksijene parçalanmasını katalizlediği saptanmıştır. Ribozomlar Ribozomların fonksiyonları bütün hücrelerde aynı olmakla beraber ökaryot ve prokaryot hücrelerde morfolojilerinde farklar vardır. Ribozomlar ya sitoplazmik matriks içinde granüller halinde veya ER zarlarının sitoplazmaya dönük yüzüne bağlı olarak bulunurlar ve GER’ mu meydana getirirler. Özellikle enzim salgılamada çok faal olan hücrelerde bol miktarda ribozom bulunur ve bunların %90 kadarı ER zarlarına bağlı olarak yer alırlar. Bu ilişki, sentezlenen proteinin, salınmak üzere, ER zarlarının boşluklarına kolayca geçerek, oradan paketlenmek için Golgi kompleksine taşındığının açık bir delilidir. Ribozomlar hücrede rastgele dağılmış olacakları gibi küçük gruplar halinde bir araya gelmiş olarak ta bulunurlar. Ribozom gruplarına polizom veya poliribozom denir. Poliribozomu teşkileden ribozom granülleri bir iplikle birbirlerine bağlı olurlar ve meydana gelen küçük ribozom zinciri yuvarlak, dalgalı, spiral şekillerde kıvrılabilir. Ribozomlar genellikle 150-250 Aº çapında az çok küresel görünüştedirler. Biri büyük, biri küçük olan iki alt birimden yapılmışlardır. ER zarlarına büyük altbirimler ile bağlıdırlar. İki altbrimin birleşmesi ile teşekkül eden tam ribozom bir monomer olarak düşünülmektedir. Ribozomların yapısının bir bütün olarak devam ettirilmesinde mağnezyum iyonları önemli rol oynar.Yüksek Mg²+ konsantrasyonunda iki ribozom birleşerek bir dimer teşkil eder. Ribozomların görevi hücrede protein sentezi yapmaktır. Genellikle ER zarları üzerinde bulunan ribozomlar hücre dışına verilicek olan proteinlerin sentezini yapar. Serbest ribozomlar ve polizomlar ise, sitoplazmada ilk protein sentezinin yapıldığı organellerdir. Buralarda sentezlenen proteinler, mevcut hyaloplazmanın artmasını –bu durum genç hücrelerin büyümeleri ve bölünüp çoğalmaları için önemlidir- ve hiyaloplazmada aktivite gösteren çeşitli katalitik enzimlerin meydana gelmelerini sağlarlar. Sentrozom Sentrozom yosun ve eğrelti gibi ilkel bitki hücrelerinde ve sinir hücreleri hariç tüm hayvan hücrelerinde çekirdeğe yakın bir yerde bulunur. Ayrıca mantar hücrelerinde de bulunur. Ancak yüksek yapılı bitki hücreleri ve yumurta gibi hücrelerde bulunmaz. Işık mikroskobunda çekirdek zarının dış tarafında açık renkli ve yuvarlak olarak görülen sitoplazma alanına sentrozom adı verilir. Sentriyol elektron mikroskobunda birbirine dikey olarak yerleşmiş içi boş silindirler şeklinde olan iki partikülden yapılmış olarak görülür. Her bir silindirin çapı 2000 A°, uzunluğ ise 3000-5000 A° kadar olur. Enine kesitte her bir sentriyol’un bir daire üzerinde paralel dizilmiş 9 adet iplikten (mikrotubulus) yapılmış olduğu görülür bu ipliklerin her biri üçer protein iplikciklerinden yapılmıştır. Asteri teşkil eden iplikler sentriyolden dışarıya doğru ışınsal olarak uzanan çok sayıda mikrotüpcüklerden meydana gelir. Bu mikrotüpcüklerın sayısı türlere göre ve hücrenin bulunduğu duruma göre farklı olur. Genellikle interfazda sayıları azalır. Fakat hücre bölünme halinde iken aster iplikleri ve uzun mikrotüpcükler halinde olur ve hatta hücre zarına temas ederler. Hücrede çift setriyol bulunur Bu çift setriyolden önce oğul setriyoller teşekkül ederek sayılarını iki çifte çıkarırlar . Herbir cift sentriyol hücre bölünürken hücrenin karşılıklı kutuplarına doğru yer değiştirir. Kutuplara daima bir çift setriyol göç eder. İğin kutuplarına göç eden sentriyollerin yapısı interfaz hücrelerindekinin aynıdır. Sentriyollerle beraber ayrılan asterler birbirinden uzaklaşmaya başlayınca aralarında iğ iplikleri teşekküle başlar. İğ iplikleri de mikro tüpcüklerden yapılmıştır. Sentriyoller kutuplara ulaştığı zaman iğ teşekkülü tamamlanır. İğ iplikleri ile sentriyollerin aynı büyüklükte mikro tüpcüklerden yapılmış olması iğin teşekkülünde sentriyol ve asterin rolünü desteklemektedir. Bununla beraber iğ teşekkülü için sentriyol ve asterin mutlaka gerekmediği de deneylerle gösterilmiştir. Sentriyolü çıkarılmış hücrelerde iğ iplikleri normal bir şekilde teşekkül etmektedir. Hatta sentriyolsüz hücrenin bir sonraki bölünmesinde de iğ iplikleri normal olarak teşekkül etmektedir. Bu sebeple ve bitkilerde de sentriyol bulunmadığı için, iğ teşekkülünde sentriyolün doğrudan bir rolü olmadığı anlaşılmaktadır. Sentriyoller belli hareket olaylarının yönetildiği merkezlerdir. Mitoz bölünmelerde iğ ipliklerini bunlar hareket ettirirler. Silyum ve flagellumla, spermatozoon kuyruğunun şekillenmelerini ve hareketlerini sağlayan da değişikliğe uğramış bir tür sentriyoldür. Silyum ve flagellumlarda bu tür sentriyolere bazal cisimcik denir. Vakuol Sitoplazmada içlerinde bir sıvı içeren boşluklara vukuol denir. Vukuol sitoplazmadan ince bir zar ile ayrılır; bu zar gerek yapı, gerekse geçirgenlik bakımından sitoplazma zarına benzer. Vukuoller özellikle gelişmiş, farklılaşmış bitki hücreleri ve Protozoa için karakteristik ve önemlidir; yüksek organizasyonlu hayvan hücrelerinde de bulunabilir. Genç ve aktif hücrelerde ufak olan veya bulunmayan vukuoller ergin hücrelerde büyük çapta, bazı bitki hücrelerinde de bütün sitoplazmanın yerini alacak, hücreyi kaplıyacak genişlikdedir, hücre sitoplazması ve çekirdek bir kenara itilmiştir. Bazı hücrelerde vukuoller hücrenin değişmez bir organeli olduğu halde diğer bazılarında gerektiğinde oluşan ve işi biter bitmez kaybolan yapılardır. Bu bakımdan vukuoller üç grup altında toplanabilir. 1.Değişmez vukuoller, 2.Luzumu halinde oluşan ve görevi bitince kaybolan vukuoller, 3.Anormal koşullarda oluşan vukuoller. 1. Değişmez vukuoller’e özellikle tatlı suda yaşayan protozoonlarda raslanır. Bir hücrelilerin kontraktil vakuolleri değişmez organellerdir. Hücrenin belirli bir bölgesinde bulunurlar ve zaman zaman büzülerek içindeki suyu dış ortama atarlar. Örneğin; Paramecium’ da daima hücrenin iki ucunda yer alan iki kontraktil vakuol şeklindedirler ve bunlar sıra ile çalışır. 2. Gerektiğinde oluşan vakuoller’ e en iyi örnek besin vakuoleridir. Bunlar protozoonlarda ve fagositoz yapan bütün diğer hücrelerde görülür. Besin vakuolu, gerek fagositoz ile ve gerekse sitostom aracılığı ile alınan besin parçalarının etrafında oluşur. Sindirim tamamlanınca besin vakuolü parçalanır ve kaybolur; yeni besin parçalarının alınması ile besin vakuolleri tekrar meydana gelir. 3. Anormal koşullarda oluşan vakuoller, hücrede çeşitli iç ve dış faktörlerin etkisi ile ortaya çıkarlar. Salgı yapan hücrelerde salgı maddeleri hücre dışına verildikten sonra meydana gelebilir. Yaşlı hücrelerde de sitoplazmanın değişmesi ile anormal vakuoller oluşur.