ORGANELLER YAPI VE FONKSİYONLARI NUKLEUS
advertisement

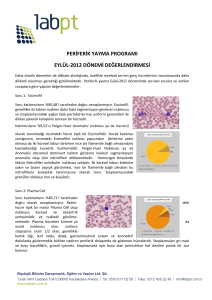
ORGANELLER YAPI VE FONKSİYONLARI Prof.Dr. Ayşegül Topal Sarıkaya İstanbul Üniversitesi Moleküler Biyoloji ve Genetik Bölümü ORGANELLER YAPI VE FONKSİYONLARI Tüm canlılarda hücreler iki tipe ayrılır. Prokaryotik ve Eukaryotik. Prokaryotik ve ökaryotik hücreler arasındaki en önemli farklılık adlandırılmalarıyla işaret edilmektedir. Prokaryot kelimesindeki “pro” Yunancada “önce” “karyon” kelimesi nukleus anlamına gelmektedir. Prokaryotik hücrede nukleus bulunmaz. Genetik materyali (DNA) nukleoid olarak adlandırılan hücrenin diğer kısımlarından bir zar yapısıyla ayrılmamış olan bölgesinde bulunur. Buna karşılık “eu” ise “gerçek” anlamında olup nukleus bir zarla çevrilmiştir. Nukleus ve zarla çevrili hücre arasında kalan kısım sitoplazma ve prokaryotik hücrelerde bulunmayan zarla çevrili özelleşmiş işlev ve yapıya sahip organellerin dışında kalan yarı katı sıvı da sitosol olarak adlandırılır. Boyut Prokaryotik 1 - 10 mikrometre çapındadır . Bakteriler prokaryotik hücrelerdir ve yaklaşık 3.5 milyar yıl önce oluşmuşlardır. Nukleus ve karmaşık organeller içermezler. Eukaryotik 10 - 100 mikrometre çapında ve daha karmaşık yapıdadırlar. Bitki, hayvan ve mantar hücreleri eukaryotiktir. İlk eukaryotik hücreler 1.5 milyar yıl önce oluşmuştur Neden organellere gereksinim duyulur Hücrelerin neden organelleri geliştirdiği hakkında birkaç neden sayılabilir. Birincisi organeller özelleşmiş işlevleri gerçekleştirebilir. İkincisi zarla çevrelenmiş organeller hücrenin bir bölümünü diğer bölümlerinden ayıran bir bölme (konteynır) gibi iş görebilir. Üçüncüsü organellerin zarları kimyasal reaksiyonların oluşması için bir bölge sağlayabilir. 1) Özelleşmiş Yapılar olarak Organeller Bu tip organele ilk örnek “sil”dir. Bu kısa filament bazı hücrelerin hareket etmesine yardımcı olmak üzere bir ayak gibi iş görebilir 2) Konteynır (bölme) olarak Organeller İnsanoğlu tarafından geliştirilen hiçbir şey bir canlı hücre kadar karmaşık değildir. Hücrede aynı zamanda birbirinden farklı yüzlerce reaksiyon olabilir. Eğer hücre tüm kimyasalları aynı karışımda içerseydi yaşaması mümkün olmazdı. Zarla çevrilmiş organeller bu kimyasal reaksiyonların gerçekleşmesi için tek tek bölmeler gibi iş görür. Bu tip organele bir örnek lizozomdur. Bu yapı sindirim enzimlerini içerir. Eğer bu enzimler hücre içinde serbest bir şekilde yüzüyor olsaydı hücre ölürdü. 3) Organel zarları kimyasal reaksiyonlar için bir alan oluşturur Bu tip organele örnek kloroplasttır. Fotosentezin ışık reaksiyonlarını yöneten moleküller kloroplastın zarında gömülü olarak bulunurlar Ökaryotik hayvan hücresi NUKLEUS Ökaryotik bir hücrenin beyni olarak ifade edilen ve kalıtsal maddenin organizasyonunun yapıldığı NUKLEUS çeşitli biyokimyasal reaksiyonların gerçekleştiği bir bölgedir. Hücrenin metabolik ve çoğalma aktivitelerinin koordinasyonu ve yönlendirilmesi için gerekli olan bir merkezdir. Hücrenin en büyük organelidir. 1700’lerin başlarında mikroskop yardımıyla canlı hücrede açık gri bir sıvının içinde (sitoplazma) daha koyu dairesel bir yapının yüzdüğü keşfedildi. 1831 de Robert Brown hücrenin merkezinde bulunan bu koyu renkli dairesel yapıyı NUKLEUS olarak tanımladı (Nukleus latince “küçük fındık”). Nukleus farklılaşmış kırmızı kan hücreleri dışında tüm ökaryotik hücrelerde bulunmaktadır. Nukleusun büyüklüğü farklı doku hücrelerinde yaklaşık 10-20 mikrometre arasında değişen bir çeşitlilik gösterebilir. Hücrelerin büyük bir çoğunluğu tek nukleusludur. Bazı patolojik durumlarda nukleus sayısının arttığı görülür. Bazen de nukleus bölünmesini sitoplazma bölünmesinin izlememesi çok nukleuslu olma sonucunu ortaya çıkarabilir. NUKLEUS Nukleus nukleus zarfı (kılıf) adı verilen hücre membranına (zarına) benzeyen bir yapıyla sitoplazmadan ayrılır. Bu yapı nukleusa ait bölmeyi belirler ve porlar içeren iki adet membrandan oluşur. İki membran arasındaki boşluk ise perinukleer aralık olarak adlandırılır. Nukleus sıvısı (nukleoplazma) sitoplazmaya benzer jel kıvamında pek çok maddenin çözündüğü bir sıvıdır. Bu maddeler nukleotit trifosfatlar, enzimler, proteinler ve transkripsiyon faktörleridir. Hücrenin genlerinin çoğu nukleusta yerleşmiştir. Nukleus DNA, RNA, nukleoplazma ve nukleolus içerir Nukleus 3 tane temel yönlendirme yapar. 1) Hücrenin en basit şekilde iki yeni hücre vermek üzere bölünmesini kapsayan hücresel çoğalması, 2) Hücrenin metabolik aktivitelerinin oluşumu sırasında hücrenin farklılaşmasını kontrol etmesi, 3) Hücrenin metabolik aktivitelerini düzenlemesidir NUKLEUS Hücrenin nukleusunda DNA ve RNA, hücreyi oluşturmak ve kontrol etmek üzere birlikte çalışırlar. DNA’daki kalıtsal bilgi RNA’ya çevrilir (transkripsiyon) ve bu ürün (haberci) nukleus porları yoluyla nukleustan dışarı protein sentezinin yapıldığı ribozoma taşınır NUKLEUS Yapısal olarak nukleus 3 ana parçadan oluşur. 1. Nukleolus, 2. Nukleus zarfı ve 3. Kromatin Nukleolus (Çekirdekçik) Nukleus sıvısı (nukleoplazma) içinde genellikle koyu bir daire şeklinde görülür. Bir veya birkaç nukleolus bulunabilir. Nukleolus ribozom alt birimlerinin birleştiği yapılardır ve ribozomları, RNA, DNA ve proteinleri içerir. Ribozomların oluşumunda etkin bir role sahiptir. Ribozoma ait ribonukleik asitin (rRNA) sentezinin yapıldığı yerdir. Nukleolusların oluşumunda nukleolus organizatörü ( nukleolusa ait organizer bölge) adı verilen özelleşmiş bölgelere sahip bazı kromozomların (nukleolar kromozom) etkisi vardır. Nukleolus etrafında zar bulunmaz. Nukleolus Aktif olarak çoğalan bir memeli hücresinde artan protein sentezi gereksinimini karşılamak için 5-10 milyon ribozom bulunur ve bunların her bölünmede sentez edilmeleri gerekir. Nukleolus denetimli ve yeterli rRNA sentezi ile ribozomal alt birimlerin oluşması için bir ribozom fabrikası gibi çalışır. Son yıllarda nukleolusun RNA modifikasyonunda ve farklı tipteki RNA’ların işlenmelerinde etkin bir rolü olduğu bulunmuştur. Nukleus zarfı Nukleus 30nm genişliğinde iç ve dış nukleus membranlar olmak üzere iki membran sisteminin oluşturduğu bir yapı ile çevrelenerek sitoplazmadan ayrılır. Dış nukleus membranı endoplazmik retikulum (ER) ile devam eder. Böylece dış ve iç nukleus zarları arasındaki boşluk doğrudan endoplazmik retikulum lümeni ile bağlanmış olur. Ek olarak dış nukleus zarı üzerinde de ER’de olduğu gibi ribozomlar yer alır. Buna karşılık iç nukleus zarı nukleusa özgü bazı proteinleri taşır. İç memranın altında nukleusa yapısal destek sağlayan fibröz bir ağ olan nukleus laminası bulunur. Nukleus laminası lamin adı verilen bir veya daha fazla fibril proteinin biraraya gelmesiyle oluşur. Nukleusa yapısal destek sağlamasına ek olarak nukleus laminası kromatinin bağlanmasında da iş görmektedir Lamin Yapısı: İki polipeptit molekülün birbirine sarılmasıyla lamin dimerleri oluşur. Dimerin baş kuyruk bağlantılarıyla doğrusal polimer oluşumu ve polimerlerin yanyana yerleşmesiyle flamentler oluşur. Nuklear Lamina’nın elektron mikrografı Diğer hücre membranlarında olduğu gibi nukleus membranları sadece küçük polar olmayan moleküllerin geçişine izin veren fosfolipit tabakalarından oluşur. Diğer moleküller fosfolipit tabakadan geçemezler. İç ve dış nukleus membranları düzenli aralıklarda birleşerek nukleus ve sitoplazma arasında taşınımı düzenleyen ve kolaylastıran 120nm çapında porları oluştururlar. Bütün bu taşınma işlemlerinin gerçekleştiği nukleus por kompleksleri nukleoporin adı verilen 50’den fazla proteinden oluşmaktadır. Genellikle nukleus transkripsiyon açısından ne kadar aktifse zarftta bulunan por sayısı o kadar fazladır. Elektron mikrosobu ile yapılan çalışmalar sonucunda por yapısını bir merkez kanalın etrafında yerleşmiş 8 adet tekerleğe benzeyen yapının (“spoke”) biraraya gelerek oluşturduğu saptanmıştır. Tekerleğe benzeyen yapılar nukleus ve sitoplazmik yüzeylerde halkalara bağlanır. Bu açıklık büyük moleküllerin geçişi için yeterlidir ve merkez taşıyıcı adı verilen bir yapı ile makromoleküllerin aktif taşınması sağlanır. Böylece porlar aracılığı ile sitosol ve nukleus arasındaki iki yönlü trafik sürekli olarak devam eder. Nukleusta görev yapan histonlar, DNA ve RNA polimerazlar, gen düzenleyici proteinler, RNA işlenmesinde görev yapan proteinler vb. Seçici bir şekilde sitosolden nukleusa taşınmaktadır. Aynı zamanda tRNA ve mRNA molekülleri de nukleusta sentezlenip sitosole taşınmaktadır. Nukleus dışına yapılan taşıma da seçici olarak gerçekleştirirlir. Örneğin mRNA’lar doğru bir şekilde işlendikten sonra taşınırlar. Ribozomal proteinler sitosolde sentezlenir ve nukleusa taşınır. Burada yeni sentezlenen rRNA molekülleriyle birleşerek ribozomların alt birimi olarak tekrar sitosole taşınır. Tekerlek-halka birliği iç ve dış membranların birleştiği noktada nukleus zarfına yerleşir. Protein flamentler sitoplazmik ve nukleus yüzeylerinden dışarı doğru uzanır ve nukleus tarafında basket filesine benzer bir yapı oluştururlar. Merkez kanal yaklaşık 40 nm çapındadır Nukleer Por Kompleksinde molekül taşınımı Küçük Moleküller por kompleksinden pasif difüzyonla kolaylıkla geçerler. Makromoleküller özellikle proteinleri sitoplazmadan nukleusa ve RNA’ları nukleustan sitoplazmaya taşıyan enerji bağımlı seçici bir taşınım ile aktarılır. Mitoz sürecinde Nukleus Hücrelerin çoğunda çekirdek zarfının bozulması mitozda profaz döneminin sonunu belirler. Nukleus bütünlüğünün bozulması mitozun yaygın bir özelliği değildir ve tüm hücrelerde gerçekleşmez. Mitoz Sürecinde Nukleus Nukleus zarfının yıkılması 3 bileşendeki değişiklik ile gerçekleşir. Nukleer zarın vesiküllere parçalanması, Nukleer por kompleksi ayrışması, Nukleer zarın altını kaplayan laminanın dağılması Nukleer laminanın dağılması filamanların lamin dimerlerine yıkılmasına neden olan laminlerin Cdc2 tarafından fosforillenmesinin sonucudur. Nukleer lamin dağılmasından sonra nükleer zar da vesiküllere parçalanır. B tipi laminler vesiküllere bağlı kalırken A ve C tipi laminler serbest olarak bulunur. Nükleere por kompleksleri de yapılarındaki birkaç por proteininin fosforillenmesiyle alt birimlerine ayrılır. İnterfaz Nukleusunun Yeniden Oluşması Mitozu başlatan Cdc2’nin inaktifleşmesi nukleus zarfının yeniden oluşmasının sinyalidir. İlk aşama nukleus yıkımda oluşan vesiküllerin kromozomların yüzeylerine tutunmasıdır. Vesiküller daha sonra kromozomların etrafında çift zar oluşturmak üzere birleşirler. Bunu nükleer por komplekslerinin yeniden yapılanması, nükleer laminanın yeniden şekillenmesi ve kromozom gevşemesi izler. Kromatin Hücrelerle ilk yapılan mikroskop çalışmaları sırasında nukleus içinde boyaları çok çabuk emen bir yapı belirlendi. Bu yapı yunancada renkli anlamına gelen “chroma” dan esinlenerek kromatin olarak adlandırıldı. Kromatin DNA ve proteinden oluşmuş bir yapıdır. Bu yapı olukça uzun ince dağınık iplikler şeklinde bulunan kromozomları meydana getirir. Koromozom Yunanca’da chroma = renkli ve soma = yapı anlamındadır. Hücre bölüneceği zaman kromozomlar kendi üzerlerinde katlanarak kalınlaşırlar ve ayrı yapılar olarak ışık mikroskobunda görülebilirler. İki tip kromatin bulunmaktadır. Ökromatin ve Heterokromatin Ökromatin DNA’nın daha az yoğun olan şeklidir ve hücre tarafından sıklıkla anlatım yapan genleri içeren DNA’dan oluşmaktadır. Heterokromatinde DNA oldukça kompakttır. Hücreler iki tip heterokromatin içerir. Sürekli Heterokromatin, hücre tarafından hiç anlatım yapmayan DNA dizilerini örneğin kromozomların telemor ve sentromerlerini oluşturan bölgeleri ve Fakültatif Heterokromatin, bazı hücre tiplerinde anlatım yapmayan fakat diğer bazı tip hücrelerde anlatım yapan DNA dizilerini içerir. Çok hücreli organizmalarda hücreler özel işlevleri gerçekleştirmek üzere özelleşirler Bu nedenle farklı gen grubları gereklidir ve anlatım yapmalıdır. Bu yüzden hücre tipleri arasında heterokromatini oluşturan DNA bölgeleri de çeşitlilik gösterir. Heterokromatin genellikle iç nukleus zarının altında bulunan özelleşmiş proteinlere bağlı şekilde bulunur. Nukleus Nukleusta morfolojileri bir diğerinden farklı olan yapılar veya bölmeler bulunur. Kajal veya kangal cisimler (küçük RNP’ler bakımından zengindir. RNP yapılanma bölgesi işlevi gördükleri düşünülmektedir) ve PML cisimler (işlevleri bilinmemektedir) en önemli 3 nukleer bölgeyi oluşturur. Organeller Yapı ve Fonksiyonları Endoplazmik Retikulum Ayşegül TOPAL SARIKAYA ENDOPLAZMİK RETİKULUM ENDOPLAZMİK RETİKULUM (ER); ÖKARYOTİK HÜCRELERDE SİTOPLAZMA İÇİNDE VARLIKLARI SADECE ELEKTRON MİKROSKOPLARI İLE BELİRLENEBİLEN TEK ZARLA ÇEVRİLİ HÜCRENİN HER BÖLGESİNDE BULUNABİLEN MEMBRAN SİSTEMİNİN BİR PARÇASI OLAN ENDOMEMBRAN KANAL SİSTEMİDİR ENDOPLAZMİK RETİKULUM İLK KEZ 1945 YILINDA BELÇİKA’DAN ALBERT CLAUDE VE ROCKEFELLER ENSTİTÜSÜ’NDEN KEİTH PORTER TARAFINDAN ELEKTRON MİKROSKOBU YOLUYLA CİVCİV EMBİRİYO HÜCRELERİNİN SİTOPLAZMASINDA MEMRANLARLA SINIRLANDIRILMIŞ KANALLARIN OLUŞTURDUĞU BU BÜYÜK AĞ SİSTEMİ GÖSTERİLDİ. PORTER TARAFINDAN BU YAPI ENDOPLASMİK RETİKULUM (ER) OLARAK ADLANDIRILDI. Endoplazmik Retikulum Daha sonra bu ağ sistemi memeli kırmızı kan hücreleri dışında tüm ökaryotik hücrelerde saptandı. Bir hücrenin toplam membranının yarısından fazlasını oluşturan ER membran sistemi hücre içinde her yöne dallanarak sitosolün her tarafına uzanan yassılaşmış keseler, dallanmış tubullerle labirent şeklinde bir ağ oluşturur. Endoplazmik Retikulum Hücreden hücreye ve hücrelerin fizyolojik etkinliğine göre kanal ve keselerin dizilişi ve miktarı değişir. Fazla protein sentezi yapan hücrelerde kanallar genişler. Bazı hücrelerde ise ince sıkı ve birbirine paralel lameller biçiminde dizilirler. ER’nin işlevleri YENİ MEMBRANLARIN OLUŞUMU İÇİN LİPİTLER, ÇOK ÇEŞİTLİ PROTEİNLER VE KARMAŞIK KARBOHİDRATLARIN SENTEZLERİNİ GERÇEKLEŞTİRİRLER. BUNUN DIŞINDA UZAMIŞ BÖLGELERİYLE DEPOLAMA GÖREVİ VE BAZI MADDELERİN HÜCRENİN BİR PARÇASINDAN DİĞERİNE TAŞINMASINDA DA İŞLEVE SAHİPTİRLER. ER MEMBRANI GENELLİKLE NUKLEUS VE GOLGİ AYGITI ARASINDA YER ALIR. BU NEDENLE NUKLEUS VE GOLGİ ARASINDA 2 MEMBRAN OLACAKTIR; DIŞ ER MEMBRANI VE NUKLEUS MEMBRANI. BUNDAN DOLAYI ER DIŞ NUKLEUS ZARININ DEVAMI OLARAK SÜREKLİLİĞİNİ SAĞLAR. ER MEMRANI ARASINDAKİ BOŞLUK LÜMEN OLARAK ADLANDIRILIR VE HÜCRE HACMİNİN YAKLAŞIK %10’NU KAPSAR İki tip ER vardır. Membranının üzerinde protein sentezinin yapıldığı ribozomları içeren Granüllü Endoplazmik Retikulum (GER) ve ribozom bulunmayan lipit sentezinden sorumlu Düz yüzlü Endoplazmik Retikulum (DER) Kas fibrillerinde bulunan ER ise sarkoplazmik retikulum olarak adlandırılır. ER İzolasyonu Düz yüzlü Endoplazmik Retikulum (DER) DER DAHA ÇOK LİPİT SENTEZİ YAPAN VE STEROİT TİPTE HORMON SALGILAYAN HÜCRELERDE (GONATLAR) YOĞUN BİR ŞEKİLDE BULUNMAKTADIR. BU NEDENLE LİPİT BİYOSENTEZİNDE ETKİLİ OLAN ÖZELLİKLE FOSFOLİPİT, YAĞ ASİDİ VE STEROİD SENTEZİNDEN SORUMLU ENZİMLERİ İÇERİR. Düz yüzlü Endoplazmik Retikulum (DER) ADRENAL KORTİKAL HÜCRELERDE KOLESTEROLÜN YAN ZİNCİRİNDEKİ PARÇALANMA MİTOKONDRİDE OLDUKTAN SONRA ÜRÜN DER’E AKTARILIR VE DAHA İLERİ DEĞİŞİKLİKLER YAPILIR. DAHA SONRA SON DEĞİŞİKLİLER İÇİN TEKRAR MİTOKONDRİYE TRANSFER EDİLİR. BU YÜZDEN İKİ ORGANEL ARASINDA STEROİD HORMON ÜRETİMİ İÇİN BİR ÇEŞİT “STEROİD VOLEYBOL” U OYNANDIĞI SÖYLENEBİLİR. Düz yüzlü Endoplazmik Retikulum (DER) DER AĞI ANAHTAR ENZİMLERİN VE BU ENZİMLERİN ÜRÜNLERİNİN DEPOLANMASI İÇİN YÜZEYİN ARTMASINI SAĞLAR. KAS HÜCRELERİNDEKİ DER’DE VESİKÜLLER VE TUBULLER, KASILMA SÜRECİ SIRASINDA AÇIĞA ÇIKAN KALSİYUMU DEPOLAYARAK İŞ GÖRÜR. KARACİĞER HÜCRESİNDE İSE GLİKOJEN DEPOLANMASI, KOLESTEROL YAPIMI VE TOKSİK MADDELERİN ETKİSİNİN ORTADAN KALDIRILMASI GİBİ ÇEŞİTLİ FONKSİYONLARA SAHİPTİR Düz yüzlü Endoplazmik Retikulum (DER) DER, GER’DE ÜRETİLEN MOLEKÜLLERİ GOLGİ AYGITI’NA VE HÜCRENİN DİĞER BÖLGELERİNE TRANSFER ETMEK İÇİN ARACI VESİKÜLLERİ DE OLUŞTURUR. KARBONHİDRAT METABOLİZMASINDA ÖNEMLİ ROL OYNAMAKTADIR. HÜCRE İÇİNDE BULUNAN BESİN MADDELERİN TAŞINMASINDA DA ETKİN BİR ROLÜ VARDIR Granüllü Endoplazmik Retikulum (GER) GER RİBOZOMLARIN VARLIĞINDAN DOLAYI ELEKTRON MİKROSKOBU ALTINDA GRANÜLLÜ BİR GÖRÜNTÜ VERMEKTEDİR. ER’YE BAĞLI RİBOZOMLAR BELLİ MEMBRAN VE ORGANEL PROTEİNLERİNİ SENTEZLER VE GEREKLİ OLDUKLARI YERLERE TAŞINMASINDA ARACI OLURLAR. UZAYAN SALGI POLİPEPTİDİ RİBOZOMDAN ÇIKAR ÇIKMAZ LÜMENDE BİRİKMEK ÜZERE GER MEMBRANINDA BULUNAN ÖZEL PROTEİNLERİN YARDIMIYLA GER MEMBRANINDAN GEÇER. Granüllü Endoplazmik Retikulum (GER) Bütün ökaryotik hücreler oldukça fazla miktarlarda GER’e sahiptir. Çünkü GER plasma membran proteinlerinin sentezi için gereklidir. GER özellikle salgı proteinleri üreten özelleşmiş hücrelerde çok fazla miktarlarda bulunmaktadır. Örneğin antikor üreten plazma hücreleri veya sindirim enzimlerini sentezleyen hücreler Granüllü Endoplazmik Retikulum (GER) GER’İN İŞLEVLERİ PROTEİN SENTEZİ, YENİ MEMBRAN PROTEİNLERİNİN ÜRETİMİ, YENİ SENTEZLENMİŞ PROTEİNLERİN DEĞİŞİME UĞRAMASI (MODİFİKASYON) PROTEİNLERİN HÜCRE İÇİNDE MEMBRANA VE DİĞER YERLEŞİM BÖLGELERİNE TAŞINMASI GER’de Protein Modifikasyonları GER ayrıca proteinlerin katlanarak üç boyutlu yapısını kazanmasından ve kalitesinin kontrolundan da sorumludur. Protein katlanması: Proteinlerin doğru bir şekilde çalışması için proteinlerin 4’cül yapılarını kazanmaları gereklidir. Disülfit bağının oluşumu ve yeniden düzenlenme: Disülfit bağları pek çok proteinin 3. 4. yapılarının kararlılığı için gereklidir. SH C H H C SH H Protein disülfit izomeraz oxidation H2 C S S CH2 + 2H+ AND 2E- H GER’de Protein Modifikasyonları Şaperon moleküller. Yeni oluşmuş proteinlere bağlanarak katlanmalarını düzenlerler. Örneğin ER içindeki Kalneksin ve Kalretikulum mitokondri içindeki proteinlerin katlanması için şaperon proteinler HSP60 ve HSP70 Kalite kontrol: Yanlış katlanmış proteinlere şaperonlar bağlanamaz ve böyle proteinler parçalanarak ortadan kaldırılır GER’de Protein Modifikasyonları Kimyasal grubların eklenmesi: metal, asetil, formil, sülfat, hidroksil grublarının eklenmesi. Örneğin Kollejen molekülünün oluşumu için lisin ve prolin bakiyelerinin hidroksilasyonu Lipitlerin eklenmesi: Hücre membranındaki lipoprotein moleküllerin oluşumu. Çok alt birimli proteinlerin oluşturulması Proteolitik parçalama: Sinyal dizilerin uzaklaştırılması Glikosilasyon: Oligosakkaritlerin proteinlere bağlanması Salgı proteinlerinin çoğu ve plasma proteinleri genellikle glikoproteinlerdir. Tek veya birden fazla olduğu durumlarda farklı şekerler özel bir sırada proteinlere eklenir. Ribozomlar Yapı ve İşlevleri Ribozomlar Yapı ve İşlevleri Ribozomlar prokaryotlar ve ökaryotlarda bulunan yapılardır. Proteinlerden ve nukleolusta yapılan prokaryotlarda 3 ökaryotlarda 4 rRNA molekülünün oluşturduğu karmaşık yapılardır. Temel görevi protein sentezi sırasında mRNA daki bilginin özgün amino asit dizisine çevirmektir. s = Svedberg units Ribozom Tipleri ve Yerleşim Bölgeleri Ribosomlar hücre içine iki şekilde bulunur : serbest ve bağlı. Elektron micrografları, ribosomları koyu granuller şeklinde gösterir. Ribosomlar hücre içinde çeşitli bölgelerde bulunmakla beraber ribozomun lokasyonu hücrenin işlevine bağlı olarak değişebilir Serbest ribosomlar Sitosolde bulunurlar Tek bir ribozom veya poliribozom ya da polisom olarak tanımlanan ribozom grubları şeklinde bulunurlar Üretilen proteinin hücre içinde kaldığı hücrelerde bağlı ribozomdan daha fazla bulunurlar Sitoplazma sıvısında bulunan proteinlerin veya önemli sitoplazmik yapısal proteinlerin sentezinden sorumludurlar Bağlı Ribozomlar ER’nin dış yüzeyinde bağlı olarak bulunurlar Üretilen proteinin salgılandığı hücrelerde serbest ribozomlardan daha fazla bulunurlar Membran yapısında yer alan proteinlerin veya sitoplazmada vesikül içinde depolanacak proteinlerin yada hücre dışına salgılanacak proteinlerin sentezinden sorumludurlar Ribosomlar ökaryotik hücrelerde mitokondri ve kloroplastlarda da bulunurlar. Sitoplazmik ribozomlardan küçüktürler Prokaryotik ve ökaryotik ribozomlar aynı kimyasal reaksiyon dizisi ile aynı işlevleri gerçekleştirirler. Ökaryotik ribozomlar çok daha büyüktür ve proteinlerinin çoğu farklıdır Mitokondri ve kloroplast ribozomları bakteri ribozomlarına benzerlik gösterir. Ribosomlar küçük fakat kompleks yapılardır Yaklaşık 20-30 nm çapında, Boyutları farklı RNA ve protein moleküllerinden oluşan büyük ve küçük alt birimlerden meydana gelir Herbir alt birim en az bir rRNA ve çok sayıda protein içerir. İki alt birim belirli sırada bir araya gelmiş 82 proteinden oluşur. Ribozomların Yapısı Prokaryotik Ribozomların bileşenleri Approximate number of nucleotides Subunit Location 16s 1,542 30s 5s 120 50s 23s 2,904 50s Type of rRNA E.coli’nin ribozomu 70s büyüklüğündedir. Yaklaşık 2/3 rRNA kalanı proteindir Ökaryotik ribozomların bileşenleri Type of rRNA Approximate number of nucleotides Subunit Location 18s 1,900 40s 5s 120 60s 5.8s 156 60s 28s 4,700 60s 80S memeli ribozomları yaklaşık eşit miktarlarda rRNA ve ribosomal proteinden oluşur. Ribozom Oluşumu Ribozomların İşlevleri