BİY 315
BİYOLOJİK ZARLAR VE
TAŞINMA
Yrd. Doç. Dr. Ebru SAATÇİ
2008-2009 Güz Yarı Dönemi
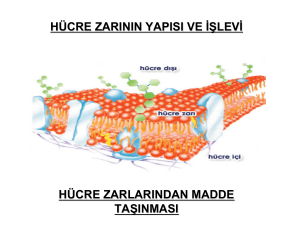
Hücre zarı ya da plazma zarı
hücrenin organellerini ve sıvı
içeriğini saran ve hücreye
yapısal bütünlük sağlayan ayrıca
çok önemli işlevleri yürüten
seçici geçirgen bir yapıdır.
Başlıca protein ve
fosfolipitlerden oluşmuş çift
katlı bir sıvıdır. Fosfolipit
tabaka membranın sıvı
bölümünü oluştururken,
fosfolipitten oluşmuş bu sıvı
tabaka içine gömülü halde
bulunan proteinler ise mozaik
bölümünü oluştururlar.
Fosfolipit tabakası
birbirine sırt sırta dayanmış
molekül gruplarından oluşur.
Kolesterol molekülleri
protein
- lipit tabakadaki
molekülleri birbirine
bağlamada görevlidir. Zar
proteinleri ise ya iyon
transferinde ya da
karbonhidratlarla
bağlanmada görev alır.
Şekilde mavi renkli yapılar
zar proteinleri olup içindeki
kanallar zarın seçici geçirgen
özelliğini belirler.
Görevleri
1) Kompartmanlaştırma; Organelleri çevreler. Bir bina içinde
bulunan farklı aktiviteler için ayrılmış odalar gibi hücre ve
organelleri de dış etmenlerden etkilenmeden fonksiyonlarını
yerine getirmek için zarla çevrilenmeleri gerekir.
2) Seçici geçirgen bariyerlerdir; maddelerin zarın bir tarafından diğer
tarafa serbest hareketini önler. Aynı zamanda bölgeler arasında
haberleşmeyi seçici olarak gerçekleştirir.
3) Solit transport yapar; az konsantre olan yerden yoğun olan yere
doğru. Şeker, aa gibi gerekli maddeleri hücre içi birikimleri
sağlanır. İyonlar içinde benzer işleri göstererek iyonik
gradientlerin meydana gelmesine yol açan bu yetenek özellikle
sinir ve kas hücreleri için önemlidir.
Görevleri
4) Dış sinyallere yanıt verir; zar yapısında bulunan reseptörler, özgül
molekülleri (ligand) ile birleşerek komplementer yapı meydana
getirirler ve sinyal iletimini başlatırlar. Farklı tip hücrelerde farklı
tip reseptörler bulunur.
5) İntersellüler etkileşim sağlar; çok hücreli bir organizmadaki
hücreler,
aralarındaki
etkileşimleri
plazma
zarlarıyla
gerçekleştirirler.
Plazma zarı, bir hücrenin komşusunu
tanımasını, yapışmasını yada madde alış- verişini sağlar.
6) Biyokimyasal aktivitelerin üzerinde yer alır; zarlar hücresel
aktiviteleri organize etmede aracı olurlar. Enzimleri multienzim
komplekslerine dönüştürülür ve tepkime düzeni kurulur.
7) Enerji iletimini sağlar; bir tip enerjinin diğer bir tip enerjiye
dönüşümünde rol alır (enerji transdüksiyonu).
Zar Lipitleri
Her bir fosfolipit tabaka,
hidrofilik (suyu seven) baş ve
hidrofobik (suyu sevmeyen)
kuyruk içerir. Hidrofilik baş
kısımları, zarın dış tabakasını
oluştururken; hidrofobik kuyruk
kısımları, diğer fosfolipit
tabakanın hidrofobik bölümüyle
karşılıklı durarak çift tabakalı
hücre mebranın iç kısımlarını
oluştururlar.
Zarda bulunan kolesterolün en
önemli görevi, yüksek ısı gibi iç
ya da dış etkenlerin zar
akışkanlığını artırmasını
engelleyerek, zar
dayanıklılığının sürdürülmesini
sağlamaktır.
Zar Hareketleri
Hücre zarı hareketsiz ve
sabit bir yapı olmayıp,
yapım ve yıkımla beraber,
kendine özgü hareketin de
sürekli olarak izlendiği özel
bir organeldir. Zar
hareketleri, her iki yana,
kendi çevresinde ve
boylamasına olabilir.
Zar Proteinleri
Her
ne kadar biyolojik zarların temel yapısı lipitler tarafından
oluşturuluyorsa da, hücrenin yaşamasıyla ilgili birçok özel fonksiyon
proteinler tarafından gerçekleştirilir. Zarda yer alan proteinlerin tipleri ve
miktarları oldukça farklıdır. Örneğin; Sinir aksonlarını kuşatan miyelin
zarlarda, zarın toplam kütlesinin %25 inden daha azı proteinken,
mitokondri, kloroplast gibi enerji üreten yapılarda bu miktar %75 kadardır.
Olağan zarlarda ise bu miktar yaklaşık % 50 dolaylarındadır.
Proteinlerin Zarlardaki Yerleşim Şekilleri
Birçok zar proteini, lipit tabakada bir taraftan diğer tarafa kadar uzanır.
Transmembran proteinler olarak adlandırılan bu proteinler amfipatik
özelliktedir. Hidrofobik kısımları lipid tabakanın hidrofobik iç kısmıyla,
hidrofilik kısımları ise lipit tabakanın her iki yanında yer alan su
tabakasıyla ilişkidedir. Zarların sitoplazmik yarısına bağlanan bazı
proteinler sadece yağ asiti zincirleri aracılığıyla lipit tabakaya tutunur.
Bazı hücre yüzey proteinleri ise küçük bir fosfolipit olan fosfatidil inositole
kovalent olarak bağlanarak lipit tabakanın dış yüzünde yer alırlar
Bunların dışındaki proteinler
ise zarın her hangi bir yüzeyinde
yer alıp, hidrofobik bölgeye
kadar uzanmazlar, zarda yer alan
diğer proteinler ile nonkovalent
bağlarla bağlanmış olarak
bulunurlar. Bunlar ekstraksiyon
metodlarıyla zarlardan çok kolay
ekstrakte edilebilirler. Bu tip
proteinlere periferal zar
proteinleri adı verilir. Bunların
aksine, transmembran
proteinler, fosfatidilinositol'e
bağlı proteinler ve yağ asitlerine
bağlı proteinler bütün lipit
tabaka deterjanlarla veya organik
çözücülerle tahrip edilmek
suretiyle elde edilebilirler. Bu
yüzden transmembran
proteinlere integral zar
proteinleri adı da verilir.
Periferal Zar Proteinleri
Yağ asitlerine Bağlanmaları:
Yağ asitlerine kovalent olarak tutunan proteinler zarda bulunan lipitlere
kovalent olarak bağlanır. Bu proteinler üç tip lipit grubuyla bağlanırlar.
Birinci tip bağlanmada (açilasyon); bir yağ asidi zinciri (miristik veya
palmitik asit) bir amid bağı aracılığıyla, sitozolik yüzde, proteinin amino
ucundaki glisine kovalent olarak bağlanır.
İkinci tip bağlanmada (prenilasyon); bir prenil grubu, bir tiyoeter bağı
ile, sitozolik yüzde, proteinin karboksil ucundaki 4. sırada yer alan
sistein'e bağlanır. Bu prenilasyonu takiben uçtaki 3 amino asit ayrılır ve
yeni oluşan karboksil uç metillenir, böylece protein zara yerleşmiş olur.
Üçüncü tip bağlanmada; proteinler, zarın dış yüzünde yer alan
glikozilfosfatidilinositol (GPI)'a bağlanır. Bu yapı genel olarak
fosfatidilinositol ve fosfatidiletanoamin içerir. Fosfatidilinositolün iki yağ asidi
zarda uzanırken, fosfatidiletanolamin bu lipit yapıyı proteine bağlar.
Fosfatidilinositol ve fosfatidiletanolamin arasında ise sayısı hücre tipine
göre değişen şeker rezidüleri (kalıntıları) yer alır.
İntegral Zar Proteinleri
Transmembran proteinlerin, polipeptit zincirinin lipit çift tabakanın
hidrofobik bölgesine gömülü olan kısmı nonpolar yan zincirli amino
asitlerden oluşurlar. Bu proteinler, lipit çift tabakayı α-sarmal (α heliks) halinde geçerler . Bu geçiş ya tek bir zincir şeklinde, tek geçişli
(single-pass), veya birkaç tane α -sarmal zincir halinde, çok geçişli
(multipass) olur.
Transmembran proteinlerin çoğu glikozillenmiştir (şeker grupları
eklenmiştir). Şeker molekülleri bu proteinlere endoplazmik retikulum
ve Golgi kompleksinde eklenir ve proteinlerin oligosakkarit zincirleri
daima ekstraselüler yüzde yer alır.
Proteinlerin sergiledikleri asimetri, sahip oldukları diğer ilave gruplar
ve çeşitli bağlar tarafından daha da artırılır. Bunlardan, sülfidril (-SH)
grupları sitoplazmik yüzde, disülfit (S-S) bağları ekstraselüler yüzde yer
alır.
Membran Proteinleri
Spektirin
Eritrositlerdeki protein moleküllerinin çoğu lipit tabakanın sitoplazmik
yüzüne tutunmuş olan periferal zar proteinleridir. Bunların içinde de en bol
bulunanı spektrin'dir. Zarın altında yer alan sitoplazma iskeletinin esas
elemanıdır.
Spektrin iki büyük polipeptit zincirinden meydana gelmiştir. Bunlar: αspektrin (~ 240 000 dalton) ve β-spektrin (~ 220 000 dalton) zincirlerdir.
Esnek yapıdaki bu zincirler birbirlerine birkaç noktada nonkovalent olarak
tutunmuştur.
Glikoforin
İnsan kırmızı kan hücre zarında
bulunan ve bütün amino asit
dizilişi tam olarak bilinen ilk
protein Glikoforin'dir. Glikoforin,
kütlesinin büyük kısmı zarın dış
yüzeyinde bulunan, 131 amino
asitten oluşmuş bir transmembran
glikoproteindir.
Glikoforin üç kısımdan meydana
gelmiştir:
1. Amino uç kısım
2. Hidrofobik orta kısım
3. Karboksil uç kısım
Band 3 Protein
Glikoforin gibi Band 3 proteini de bir transmembran proteindir.
Glikoforinden farklı olarak çok geçişli (multipass) bir zar proteinidir.
Yaklaşık 900 amino asitten oluşmuştur.
Eritrositlerdeki karbondioksitin oksijen ile değişimi bir anyon
değiştirici transport proteini: Band 3 proteini tarafından yapılır. Band 3
proteinleri dondurma-kırma elektron mikroskobi teknikleriyle
incelendiğinde zar içi partiküller olarak görülür.
Bakteriorodopsin
Bir bakteri olan Halobacterium halobium'un zarı üzerinde yer alan çok geçişli bir zar
transport proteinidir. Bu protein bakterinin zarında aktif proton (H+) pompası olarak
görev yapar. Zarda, birbirine oldukça yakın olarak yer alan 7 α-sarmal halinde kıvrılmış
olarak, lipit çift tabakada yer almaktadır. Her bir α -sarmal yaklaşık 25 amino asitten
meydana gelir. Bu α -sarmal segmentler zarın her iki yanında hidrofilik segmentlerle
birbirlerine bağlanırlar.
Zar Proteinlerinin Saf Olarak Elde Edilmesi
Genel olarak transmembran proteinler, lipit tabakayı tahrip ederek
hidrofobik ilişkileri bozan çeşitli ajanlar tarafından çözündürülebilir.
Bu ajanlar içinde en fazla kullanılan deterjanlardır (SDS; gibi).
Deterjanlar misel şeklinde toplanma eğilimi gösteren amfipatik
moleküllerdir. Zarlarla karıştırıldıklarında deterjanların hidrofobik
uçları zar proteinlerinin hidrofobik bölgelerine bağlanır, böylece lipit
moleküllerin yerini değiştirirler. Deterjan molekülünün diğer ucu
polar olduğundan, protein molekülünün lipit tabakadan ayrılarak bir
protein
- deterjan kompleksi halinde solüsyon içerisine geçmesini
sağlar. Bu komplekse nadiren çok kuvvetli bağlanmış lipit
molekülleri de katılabilir.
Deterjanlar dializ veya herhangi bir kromatografi tekniği
kullanılarak uzaklaştırılmadan önce ortama fosfolipidler karıştırılır.
Böylece sadece o proteini ihtiva eden özel bir fosfolipit çift tabaka
elde edilir ve o proteinin istenen fonksiyonu bu ortamda araştırılır.
Deterjanların polar uçları, ya sodyum dodesil sülfat (sodium dodecyl
sulfate) (SDS) ta olduğu gibi elektiriksel olarak yüklü (iyonik) ya da
triton'da olduğu gibi yüksüzdür (aniyonik).
Zar Proteinlerinin Eritrositlerde İncelenmesi
Ökaryotik hücrelerde hücre zarı ile yapılan çalışmalarda çoğunlukla kırmızı
kan hücreleri kullanılır, çünkü:
1. Diğer hücrelerle karışma tehlikesi olmaksızın bol olarak elde edilebilir.
2. Çekirdek veya diğer organellere sahip olmadığından diğer iç zarlarla
karışmadan, doğrudan saf olarak eritrosit zarı elde edilebilir.
3. Boş kırmızı kan hücre zarları veya kalıntıları (ghost) elde etmek çok
kolaydır. Kan hücresinin sitoplazmasından daha düşük konsantrasyonlarda
(hipotonik) solüsyonlar kullanıldığında, normal difüzyon kurallarına göre
dış ortamdaki sıvı hücre içine girerek kırmızı kan hücrelerinin patlamasına
(liziz) neden olur.
4. Zar kalıntıları ile fazla parçalanmadan, sadece tek noktadan yırtıkken
çalışılabilir (böylece, çalışmada kullanılacak solüsyonlar isteğe göre zarın
her iki yüzüyle temas edecektir), veya yırtığın kapanması sağlanır. Bu
özellikten faydalanılıp, kullanılan değişik iyonik konsantrasyonlar sayesinde
iç yüzü dışa gelmiş veziküller elde edilebilir.
Bu tür çalışmalarla bazı zar proteinlerinin lipit tabakayı boydan boya
geçtiği ve lipit çift tabakanın lipit kompozisyonunun birbirinden farklı
olduğu gösterilebilinmiştir. İnsan eritrosit zarlarında, SDS
poliakrilamit jel elektroforezi ile yapılan çalışmalarla, molekül ağırlığı
15000- 250000 arasında değişen 15 büyük protein bandı elde
edilmiştir Bu proteinlerden üçü, spektrin, glikoforin ve Band 3
toplam proteinlerin % 60 ı kadardır.
Transport Sistemleri
Biyolojik membranlarda
madde geçişini sağlayan
proteinler transport
sistemlerini oluşturur.
Transport sistemleri,
fonksiyonel açıdan hareket
eden moleküllerin sayısına ve
hareketin yönüne ya da
hareketin dengeye doğru veya
dengeden uzak olmasına göre
tanımlanabilirler. Hareket
eden moleküllerin sayısına ve
hareketin yönüne göre
transport sistemleri, üniport ve
kotransport olmak üzere iki
tiptir.
Kotransport sistemler, simport ve antiport olmak üzere iki tiptir: 1)
Simport sistem, iki farklı molekülü aynı yönde hareket ettiren sistemdir.
Bakterilerdeki proton ve şeker transportörleri, memeli hücrelerindeki Na+
ve şeker (glukoz, mannoz, galaktoz, ksiloz, arabinoz) trasportörleri, Na+ ve
amino asit transportörleri simport transportörlerdir. 2) Antiport sistem,
iki farklı molekülü zıt yönde hareket ettiren sistemdir. Na+ iyonu hücre
içine girerken Ca2+ iyonunun hücre dışına çıkması antiport sistem örneğidir.
Özellikle uzun zincirli yağ asitlerinin ve NADH’in mitokondriye
alınmasında antiport sistemler önemli rol oynar:
Kolaylaştırılmış Difüzyon
Kolaylaştırılmış
diffüzyon ile
membranlardan geçiş
başlıca üç şekilde
olabilir: 1) Membranı
tümüyle kateden ve
porinler diye
adlandırılan kanal
proteinleri, 2) Molekül
ağırlığı ve tipine göre
farklı olan taşıyıcı
proteinler, 3)
Proteinlerin flip-flop
hareketi ile
gerçekleştirilir.
“Pin” (flip) durumunda boşalmış olan
taşıyıcı, sonraki döngüyü tamamlamak
için orijinal konformasyonu olan “pon”
(flop) durumuna döner.
Aktif Transport
Aktif transport, moleküllerin termodinamik dengeden uzağa yani
konsantrasyon veya elektrokimyasal gradientin zıt yönünde
taşınmaları açısından diffüzyondan farklıdır; bundan dolayı
enerji gerektirir. Aktif transport için gereken enerji, ATP’nin
hidrolizinden, elektron hareketinden veya ışıktan sağlanabilir.
Aktif transport ile biyolojik sistemlerde elektrokimyasal
gradientlerin devamlılığı sağlanır. Genellikle hücreler, hücre içi
ortamda net bir negatif elektriksel potansiyel ile bir arada düşük
bir intrasellüler Na+ konsantrasyonu ve yüksek bir intrasellüler
K+ konsantrasyonu sağlarlar.
İntrasellüler ve ekstrasellüler arasında gardientleri sağlayan
Na+/K+ ATPaz pompasıdır. Na+/K+ ATPaz pompası,
membrana bağımlı Na+/K+ ATPaz tarafından her ATP
molekülünün ADP’ye hidrolizi sırasında 3Na+ iyonunun hücre
içinden dışarıya ve 2K+ iyonunun dışarıdan hücre içine
hareketini sağlar.
Kolaylaştırılmış diffüzyon ve aktif transport pek çok özelliği
paylaşırlar; benzerliğin söz konusu olduğu noktalar şöyledir: 1)
Solüt için spesifik bir bağlanma bölgesi vardır. 2) Taşıyıcı, satüre
olabilir; bundan dolayı bir maksimum transport hızına (Vmax)
sahiptir. 3) Solüt için bir bağlama sabitesi (Km) vardır ve sonuçta
tüm sistemin bir Km’si vardır. 4) Yapısal olarak benzer olan
yarışmalı inhibitörler transportu bloke ederler.
Kolaylaştırılmış diffüzyon ile aktif transport arasındaki temel
farklar şunlardır: 1) Kolaylaştırılmış diffüzyon iki yönlü işlerlik
gösterebilir, halbuki aktif transport genelde tek yönlüdür. 2) Aktif
transport, her zaman bir elektriksel veya kimyasal gradiente karşı
meydana gelir ve bundan dolayı enerji gerektirir.