DNA Hataları ve Düzeltme Mekanizmaları
advertisement

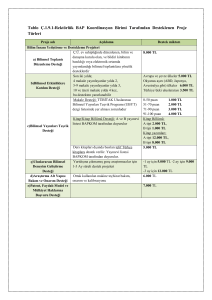
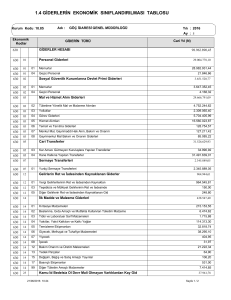
DNA Hataları ve Düzeltme Mekanizmaları Bir canlıya ait tüm genetik bilgiyi taşıyan DNA molekülü doğal olarak veya çevresel faktörlerin etkisiyle sürekli hasara maruz kalmaktadır. İnsan hücrelerinde metabolik aktiviteler ve çevresel faktörler (UV ışığı gibi) sonucu günde 1 milyon hücrenin zarar görmesi olasıdır. Bu etkenler, DNA'nın yapısını ve dahası diğer nesillere aktarılan genetik bilgiyi değiştirebilirler. Bu değişimler yararlı olabileceği gibi, ölümcül sonuçlara neden olabilecek kadar da zararlı olabilir. Bu yüzden, bütün canlı hücreleri, evrim süreçleri boyunca nesillere değişmeden aktarılması gereken DNA molekülünü koruma mekanizmaları geliştirmişlerdir. Küçük hasarlar çoğunlukla DNA onarım sistemleri tarafından onarılır. Yüksek düzeydeki hasarlar apoptozisi uyararak "hücre ölümüne" yol açar. Böylelikle organizma kendini korumuş olur. Orta derecedeki hasarların birikimi ise mutasyonlara neden olur. Hücre tüm bu DNA hasarlarına farklı metabolik yollar ile cevap verir. Ağır DNA hasarları hücrenin apoptozis yolunu aktive ederek hücreyi ölüme götürür. Hücre, DNA hasarlarını "DNA tamir mekanizmaları" ile tamir edebilir. DNA hasarı ikileşme sırasında tamir edilemezse mutasyona ve sonuç olarak genomik kararsızlığa, kanser ve yaşlanmaya neden olur. DNA tamir sisteminde 100’den fazla gen rol oynar ve bu genlerin kodladığı proteinler tamir mekanizmalarında görev alırlar. Her bir insan hücresinin DNA'sında günde yaklaşık olarak 104 adet kodlanmayan veya yanlış kodlamaya neden olan hasar meydana gelmektedir. Mitokondrial DNA'da nokta mutasyonlarının birikiminin yaşa bağlı olarak arttığı, bu nedenle özellikle mitokondrideki oksidalif DNA hasarının yaşlanma ile ilişkili olduğu düşünülmektedir. DNA tamir mekanizmaları genomik kararlılığın devamını sağlayan sistemlerdir. Mutasyon DNA dizileri, normal olarak replikasyon işlemi sırasında tam olarak kopyalanır. Ancak DNA replikasyonu ve onarım sırasında, nadiren yeni sekanslar oluşması ya da hatalar meydana gelir. Bu hatalara mutasyonlar denir. Mutasyonlar evrimin, dolayısıyla değişimin ve yenilenmenin kaynağıdır. Mutasyonlar, tek bir nükleotitte gerçekleşiyorsa buna nokta mutasyon ya da daha fazla sayıda nükleotidini etkiliyorsa bölgesel mutasyon (segmental mutasyon) adını alır. Mutasyonlarda kendi içinde oluş şekline, değişiklik türüne göre sınıflandırılabilir. Buna göre: (1) Yer değiştirme (Substitution mutation) mutasyonları, bir nukleotidin diğeri ile yer değiştirmesi, (2) Rekombinasyonlar (Recombination), bir dizinin diğer bir sıra ile değişimi, (3) Silme (Deletions), DNA,’dan bir ya da daha çok nükleotidin çıkarılması, (4) Eklenme (Insertions) çift kollu bir DNA segmenti ve dizisine bir ya da daha çok nükleotidin eklenmesi, (5) Dönme (Inversions), çift iplikli DNA sarmalının iplikçiklerinden birinin yada ikisinin 180° dönmesi şeklinde olur. 1.Yer Değiştirme Mutasyonları (Substitution Mutation) Yer değiştirme mutasyonları ikiye ayrılır. Bunlardan ilki geçiş tipi mutasyonları (transizyon tipi mutasyonlar) ve diğeri çapraz yer değiştirme mutasyonları (transversiyon tipi mutasyonlar) olarak ayrılır. Transizyon tipi mutasyonlarda bir pürin, bir pürinle; bir primidin bir primidinle yer değiştirir. Yani değişikler A ve G (pürinler) arasında, ya da C ve T (pirimidinler) arasındadır. Çapraz yer değiştirme mutasyonları olan transversiyon tipi mutasyanlarda bir purin, bir pirimidin ile yer değişmektedir. Transizyon tipinde dört tip değişim görülür. A-G ‘nine, G-A’nine değişir. T-C’e ve elbette C-T’e değişebilir. Transversiyonda ise sekiz tip yer değiştirme gözlenir. Bunlar; G-A, A-G, C-T, T-C, C-G, G-C, A-C, C-A, T-A, A-T, G-T ve T-G’dir. Bu mutasyonlarla protein kodlama bölgelerinde meydana gelen değişiklikler, çeviri sonucu ortaya çıkacak protein ürünün yapısını ve/veya görevini etkilerler. Buna bağlı olarak da bu mutasyonları sınıflandırabiliriz. Oluşan yer değiştirme mutasyonu eğer aynı amino asitin üretimine neden oluyorsa bu mutasyonlar proteinin yapısı ve görevini etkilemez dolayısıyla sessiz (eşanlamlı) mutasyonlar adını alır Eğer proteinin yapısına ve/veya görevine etki ederse, bunlar anlamlı (farklı anlamlı) mutasyonlar adını alırlar. Burada değişiklik merkezde olan önemli bir amino asitte değilse proteinin 2.cil ve 3.cül katlanmalarını etkilemez. Bununla beraber merkezi bölgede yer alan aminoasitlerden birinde ise proteinde ciddi bozukluklara yol açar. 2.Rekombinasyon Rekombinasyonel mutasyonlarda iki tip rekombinasyon bulunur. Bunlardan ilki karşılıklı parça değişimini içeren krosing-over (Crossing over -Reciprocal recombination), diğeri de karşılığı olmayan, gen değişimlerini (Gene conversion-nonreciprocal recombination) içerir. Karşılıklı rekombinasyon, homolog kromozomlar arasındaki homolog sekanslarında, bunlar arasında yer alan yeni dizilerin de değişimini içerir. Karşılıklı olmayan, gen değişimlerini içeren rekombinasyonda ise kromozomda yer alan bir dizinin yer değiştirmesi değil genellikle kaybedilmesi ile ortaya çıkar. 3.Silme ve Eklenme DNA’da oluşan mutasyonların bir diğer tipi de insersiyon(ekleme) ve delesyon(çıkarma) mutasyonlarıdır. Silinmeler ve eklenmeler, çeşitli mekanizmalarla oluşabilir. Bu mekanizmalardan ilki, eşit olmayan krosingover(unequal crossing over) oluşmasıdır. Burada gerçekleşen herhangi bir kromozomun, bir DNA parçasının homoloğundaki diğer karşılıklığı ile yer değiştirmesi sırasında, bir eksilir veya silinirse, iki kromozom arasındaki eşitlik, benzerlik sona erer. Birbiri ardına(Tandem) gelen DNA segmentleri arasında oluşan kayma nedeniyle ve büyük olasılıkla eşit olmayan bir çaprazlama gerçekleşir. Başka bir mekanizmada intrastrand(şerit-arası slime), DNA ipliği içinde silinme mekanizmasıdır. Bu bir çeşit bölgeye özel rekombinasyondur. Kromozom üzerinde yer alan, tekrar eden diziler yine krosingover sırasında kromozomlar arasında yer değiştirirler. Bu şekilde aynı kromatidler üzerinde yer alan ve tekrarlayan diziler arasında ve bu dizilere özgün yeni bir düzenleme, rekombinasyon oluşur ve diğer bir deyişle intrakromozomal(Kromozomlararası)geçişler ortaya çıkar. Üçüncü bir mekanizma çoğaltma aşamasında ortaya çıkan kaymalar (replication slippage)veya kayma esnasında iplikçik kayıpları(Slipped-strand mispairing)’dır. Bu olaylar kısa tekrar eden DNA dizileri olan bölgelerde meydana gelir. Böyle bir mutasyon, çerçeve kayması mutasyonu (Frameshiftmutation) olarak bilinir. Bunun sonucunda indeller bir veya daha çok sayıda aminoasit değişikliğine yol açabilirler. Bunun sonucunda da oluşacak protein determinal uzama kodonu yok olabilir veya anormal uzunlukta bir protein şekillenebilir veya yeni bir durdurma kodonu meydana getirebilir. 4.İnversiyon DNA’nın yeniden düzenlenmesini sağlayan birbirine zıt iki aşamalı bir işleyiştir. Buna gore: (1) Kromozom üzerinde bir kırılma ve katılım olur veya (2) Krosingover sırasında birbirine homolog kromozom bölgeleri arasında kopmalardan sonra eklenme aşamasında zıt yönlü bağlanmalarla ortaya çıkar. Bilinenin versiyonların büyük bir çoğunluğu, genellikle, yüzlerce veya binlerce nükleotit uzunluğunda DNA parçalarını içerir. Mutasyonlar, replikasyon nedeniyle eklenmiş yanlış bazlar yanı sıra hidrolitik reaksiyonlar, metilasyonlar, oksidatif solunumunun ve çevresel toksinleri içeren redoks döngüsel olaylarının ürünü reaktif oksijen bileşikleri ile de DNA hasarı oluşur. Reaktif oksijen bileşikleri enflamasyon alanlarında bağışıklık sistem hücreleri (makrofajar ve nötrofiller) tarafından da üretilir. Endojen nedenler ile oluşan oksidayon ile DNA hasarı oranı hücre başına yaklaşık 10.000 olarak saptanmıştır. Reaktif oksijen türleri DNA’da baz kaybına, yanlış eşleşmeye veya iplik kırıklarına yol açabilir. Bu hasarların içinde en az oluşanı ama en zor onarılanı çift iplik kırıklarıdır. Çevresel DNA hasar ajanlarından en önemlisi ise ultraviyole ışınlarıdır (UV). Ozon tabakası, güneş UV spektrumunun en zararlı kısmını (UV-C) absorbe eder ancak kuvvetli güneş ışığına bir saat maruz kalan her hücrede ozon tabakasından geçen UV-A ve UV-B yaklaşık 100.000 lezyon oluşmasına sebep olur. İyonize radyasyon (IR) da değişik formlarda DNA hasarı oluşturabilir. Bazı IR doğal bulunan radyoaktif bileşiklerden kaynaklanır. Örneğin uranyum radyoaktif radon gazı oluşturarak akciğer kanseri riskinin artmasına yol açmaktadır. Günümüzde kansere sebep olan en önemli çevresel etkenler birisi de sigaradır. Aflatoksin içeren besinler, özellikle şarkuteri ürünlerinde koruyucu olarak kullanılan nitritler, türevleri ve aşırı pişirilmiş et ile açığa çıkan kimyasallar DNA hasarına yol açan diğer önemli dış etkenler arasında sayılabilir. DNA ONARIM MEKANİZMALARI DNA onarım hatalarının, genomik kararsızlıkla karakterize sendromlara ve kanser insidensinde artışa varan sonuçlar doğurduğu düşünüldüğünde, DNA onarımının önemi anlaşılmaktadır. İnsanda DNA onarımı ile bağlantılı 130 genin klonlanması ve dizi analizi tamamlanmıştır. DNA onarım genleri iki alt gruba ayrılabilir: 1) DNA onarımında sinyal iletimi ve onarımın düzenlenmesi ile ilgili genler 2) Hatalı eşleşme onarımı, baz çıkarma onarımı, nükleotit çıkarma onarımı gibi ayrı onarım mekanizmaları ile ilgili genler a. DNA Çift-Zincir Kırığı Onarımı DNA çift zincir kırığının kaynakları: • İyonize radyasyon • Topoizomeraz inhibitörleri (etoposide, adriamycin) • V(D)J rekombinasyonu DNA çift zincir kırıkları (DSBs), DNA hasarının en yıkıcı şeklidir. Onarılmazsa kromozomların kırılmasına ve hücre ölümüne varan sonuçlar doğurabilir. Yanlış onarılırsa kromozom translokasyonuna ve kansere sebep olur. DSBs’ye neden olan en önemli eksojen ajan iyonize radyasyondur. Ayrıca radon bozunumu ve antikanser ilaçlar da etkilidir. Oksidatif serbest radikaller oluşturan Bleomisin, Adriyamisin, Etoposit topoizomeraz II’yi inhibe ederek protein köprülü DSBs’ler meydana getirirler. DSBs oluşturan endojen ajanlar ise serbest radikaller oluşturan oksidatif metabolizma ve V(D)J rekombinasyonudur. V(D)J rekombinasyonu B ve T lenfositlerini kodlayan genlerin düzenlenmesi esnasında DSBs oluşturur. DSBs ‘in onarımı için 2 yolak vardır. a.1. Homolog Rekombinasyon DNA çift zincir kırıkları, genetik bilgi korunarak, homolog DNA ile rekombinasyon aracılığıyla tamir edilir. Mayada bu yol çift zincir kırığı onarımında baskın olarak kullanılır. İnsanda homolog olmayan uç bağlanması ile eşit önemdedir. a.2. Homolog olmayan uçların bağlanması Homolog bir kromozomdan faydalanmaksızın DNA uçlarının bağlanmasının biyokimyasal bir yoludur. Çünkü kırık DNA uçları bağlanabilir durumda (ligatable) olmayabilir ve bu yol bazen genetik bilgide kayba da neden olur. Homolog olmayan uç bağlanmasındaki hatalar iyonize radyasyon duyarlılığına ve immün yetersizliğe (severe combined immunodeficiency) neden olur. X ışınları ve peroksitler gibi bazı kimyasallar DNA omurgasında kırıklara neden olurlar. Tek zincirdeki basit kırıklar DNA ligaz tarafından onarılır. Ancak, DNA ligaz, sadece, 5’-fosfat ve 3’- hidroksil gruplarına sahip uçları birleştirebilir. a.3. Oksidatif Hasarın Onarımı Elektron taşıma sisteminin mitokondride yer alması ve mRNA sentezi esnasında RNA polimerazın bir hata okuma (proofreading) mekanizmasına sahip olmaması nedeniyle, oksidatif hasar, mitokondride kromozomal DNA’ya göre daha sıklıkla meydana gelir. Mitokondriyal solunum sonucu oluşan serbest radikaller (-O2 / Süperoksit, H2O2 / Hidrojen peroksit, -OH / Hidroksil), tek zincir kırıklarına ve 8-oksoguanin ve timin glikol gibi hasarlı bazların oluşmasına sebep olur, hücreler bu tip hasarlı bazların onarımı için özel glikozilazlar içerir. b. Baz Hasarı Onarım Mekanizmaları b.1. Hasarın Doğrudan Geri Döndürülmesi (Direct Reversal) Hasarın geri döndürülmesi onarım için en kolay yol gibi görünmesine karşın çoğu durumda termodinamik ve kinetik nedenlerden dolayı reaksiyonun geri dönmesi mümkün değildir. Bazı durumlarda ise enzim aracılığı (Fotoliyaz ve O-6-Metil-DNA-alkiltransferaz) ile gerçekleşen tek adımlı reaksiyonlar ile hasar onarılır. Siklobütan pirimidin dimerleri (CPDs), fotoliyaz enzimi tarafından ayrılarak hasar giderilir. Reaksiyona fotoreaktivasyon denir. CPD fotoliyazlar bakterilerde, mantarlarda, bitkilerde ve çoğu omurgalıda bulunur ancak plasentalı memelilerde bulunmaz. b.2. Baz Çıkarma Onarımı (Base Excision Repair/BER) DNA hasarının doğrudan geri döndürülmesinde, bazlardaki her kimyasal değişiklik kendine özgü bir onarım mekanizması gerektirir. Ancak, hücreler birçok kimyasal hasar tipini düzeltebilecek genel bir onarım mekanizmasına ihtiyaç duyarlar. Bu da eksizyon onarımdır (excision repair). Yanlış yerleştirilen ve hasarlı bazları uzaklaştırmak için kullanılan onarım mekanizmasıdır. Her yanlış baz tipine özgün birçok yolak vardır. Bu yolaklar 2. ve 3. basamaklar ortak olmak üzere 3 adımdan oluşur: 1. Yanlış bazın uygun bir DNA N-glikozilaz tarafından uzaklaştırılması ve bir AP (Apürinik / Apirimidinik) bölge oluşması. AP bölgeleri spontan olarak kaybolan ya da glikozilaz etkisiyle uzaklaştırılan DNA bölgeleridir. Bir memeli hücresi günde 10000 pürin ve 500 pirimidin kaybeder. 2. Hasarlı DNA’ya AP bölgesinin 5' ucuna doğru AP endonükleaz tarafından çentik atılması ve AP bölgesine komşu bir 3'-OH ucu oluşturulması. 3. AP bölgesinin kesilip çıkarılarak (excision) uzaklaştırılması ve DNA polimeraz tarafından 3'-OH ucunun uzatılması. İnsan hücrelerinde çok sayıda DNA N-gikozilaz tanımlanmıştır. Diğer ökaryotik organizmalarda ve prokaryotlarda da benzer yapıda glikozilaz enzimi bulunmaktadır. DNA N-gikozilaz, DNA sarmalı üzerinde hatalı eşleşmeden kaynaklanan bükülmüş yapıyı tanır, baz ve deoksiriboz arasındaki N-glikozidik bağı hidroliz ederler. Ayrıca glikozilazlar bazların yüksek afinite gösterdiği bağlanma bölgelerine sahiptirler. Bu iki etken birleşince yanlış eşleşen bazın DNA çift sarmalından çıkarılması kolaylaşır. Urasil DNA N-gikozilazın aktivitesi görülmektedir. Spontan deaminasyon hatalı baz eşleşmesine ve böylece DNA replikasyonu esnasında kalıcı hale gelen mutasyonlara sebep olur. Örneğin; deaminasyon sitozini urasile çevirir ve sonrasında, urasil replikasyon esnasında adenin ile eşleşir. Urasil N-glikozilaz hatayı tanır ve urasili çıkarır. DNA N-gikozilazlar ayrıca AP liyaz aktivitesine sahiptirler. Bu şekilde AP bölgedeki 3'-OH ucunda DNA omurgasını keserler. Bir sonraki adımda AP endonükleazları 5’ fosfodiester bağını hidroliz ederler ve uygun nüklotidin yer alması için abazik deoksiribozu uzaklaştırırlar. Son olarak, DNA polimeraz (polimeraz-β) tarafından doğru nükleotidin yerleştirilmesi ve zincirin ligasyonu ile onarım tamamlanır. b.3. Nükleotid Çıkarma Onarımı (Nucleotide Excision Repair / NER) DNA bazları üzerinde büyük eklentiler oluşturan birçok çeşit hasarı tanıyabilen bir onarım mekanizmasıdır. Mikoplazmadan memelilere kadar geniş bir yelpazedeki organizmalar tarafından kullanıldığı belirlenmiştir. Birçok DNA hasarının özellikle de heliks distorsiyonuna neden olanların onarımında etkindir. İnsanlarda güneşten gelen UV ışığının karsinojenik etkilerine(dimerler) ve sisplatin, 4nitrokuinolin oksit gibi etkenlerle reaksiyon sonucu oluşan büyük eklentili hasarlara karşı önemli bir savunma mekanizmasıdır. NER mekanizmasının anahtarı; 1. Hasarın tanınması 2. Protein kompleksinin hasarlı bölgeye bağlanması 3. ~24-32 nükleotid uzunluğunda bir fragment içinde bırakacak şekilde lezyonun her iki tarafından hasarlı zincirin kesilmesi (incision) 4. Degradasyon (hasarı içeren oligonükleotidin uzaklaştırılması) 5. DNA sarmalı üzerinde meydana gelen boşluğun DNA polimeraz tarafından doldurulması (polimerizasyon) 6. Ligasyon Kalıtsal sendromları (Xeroderma pigmentosum/XP, Cockayne syndrome/CS, Trichothiodystrophy/TTD) olan bireylerde NER mekanizmasında bozukluklar saptanmıştır. Bu bireylerde güneşe duyarlılık, bazı dokularda erken yaşlanma, nörolojik bozukluklar ve genellikle UV kaynaklı cilt kanseri insidensinde artış gözlenir. Transkripsiyona Bağlı Onarım (Transcription Coupled Repair/TCR): Genomun her bölgesi eşit etkinlikte onarılmaz, genin transkripsiyona uğrayan zinciri aynı genin transkripsiyona uğramayan zincirine göre daha etkin olarak onarılır. Transkripsiyona bağlı onarım, RNA polimeraz II (mRNA’yı sentezleyen enzim) bir DNA lezyonu ile karşılaştığında aktive olan DNA onarımıdır. Global genomik onarım (global genomic repair), transkripsiyondan bağımsız olan DNA onarımıdır. Transkripsiyona uğrayan gen bölgelerindeki timin glikollerinin, genomun herhangi bir yerindeki timin glikollerinden daha hızlı tamir edildikleri gösterilmiştir. Transkripsiyona bağımlı bu özellik, baz çıkarma onarımında (TC-BER) ve hatalı eşleşme onarımında da görülür. b.4. Hatalı Eşleşme Onarımı (Mismatch Repair) Bu onarım mekanizması, DNA replikasyonu esnasında meydana gelen ve çift sarmalda anormal boyutlara neden olan, normal bazların hatalı eşleşmesi şeklindeki hataları düzeltir. Örneğin, E. coli’de hatalı eşleşme 7 proteinden oluşan bir sistem tarafından belirlenir. Bu proteinler, mutS, mutL, mutH, uvrD, ekzonükleaz I, SSB ve DNA polimeraz III tür. E. Coli DNA’sında, (5')GATC dizisindeki adeninler özel bir metilaz olan “Dam Metilaz” tarafından metillenmiştir. Replikasyon esnasında kalıp zincir metillenmiş durumdadır. Ancak, yeni sentezlenen zincir birkaç dakikalık bir gecikme ile metillenir. Bu zaman sürecinde yeni zincirdeki hatalı eşleşen bazlar mutS tarafından tanınır. Sırayla mutL ve mutH bir kompleks oluşturmak üzere sisteme katılırlar ve DNA boyunca çift yönlü olarak metilenmemiş bir GATC buluncaya kadar hareket ederler. MutH’deki endonükleaz fonksiyonu metil grubunun karşısında metillenmemiş zincire bir çentik atmak üzere aktive olur. Metillenmemiş zincir, ekzonükleaz I, SSB ve uvrD helikaz’ın birlikte hareketi ile uzaklaştırılır. DNA polimeraz III doğru DNA zincirini tekrar oluşturur ve ligasyon ile onarım sona erer. GATC bölgesi ile hatalı eşleşme arasındaki uzaklık en çok 1000 bç olabilir. Bu nedenle hatalı eşleşme onarımı etkili bir onarım mekanizması değildir. Ökaryotlar da E.coli‘deki mut proteinlerine homolog proteinlere sahiptir. Ancak yeni sentezlenen zinciri ayırmak için E. coli’ye özgü metilasyon işleminin yerine kullanılan mekanizma henüz tam olarak anlaşılamamıştır. Hatalı eşleşme onarım mekanizması genlerinde mutasyon olan bireylerin kalıtsal nonpolipozal kolon kanserine (HPNCC) yatkın oldukları tespit edilmiştir SONUÇLAR Hücreler DNA hasarının onarımı için birçok mekanizmaya sahiptir. Birçok DNA onarım geni, DNA metabolizmasında başka görevler de üstlenir. Örneğin, TFIIH, hem transkripsiyonun başlangıcında, hem de nükleotid çıkarma onarımında görev alır. Çift zincir kırığı onarım genleri de ayrıca immün sistemin oluşumunda görev alır. DNA onarımındaki bozukluklar, meme, kolon ve cilt kanseri gibi birçok kanser türüne neden olduğu gibi, ayrıca, büyüme ve beyin anomalilerine de sebep olur. Birçok kanser hücresinde, artırılmış DNA onarımı, kanser tedavisine gelişen dirençle ilişkilendirilebilir, bu nedenle, tedavi şekli olarak, DNA onarımında görev alan proteinlerin inaktivasyonu veya ekspresyonlarının azaltılması yönüne gidilebilir. Kaynaklar: 1- YRD.DOÇ.Dr.Yosun MATER / MOLEKÜLER EVRİM DERS NOTLARI 2-MUTATION, DNA DAMAGE, REPAIR MECHANISMS AND THE RELATION OF CANCER / Bilge DEBELEÇ-BÜTÜNER , Gülten KANTARCI