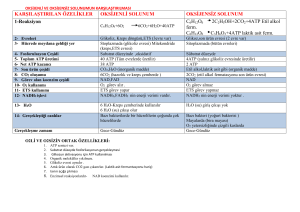
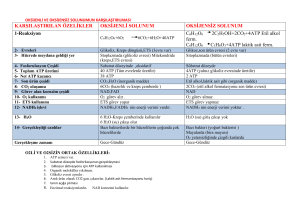
Glukoz - SABİS
advertisement

Karbohidrat Metabolizması Giriş Karbonhidratlar, önemli gıda maddelerinin en başında yer alır. Yaklaşık olarak günlük enerji ihtiyacının % 50 si karbonhidratlarla karşılanır. Yetişkinde günlük enerji gereksiniminin 2400 kcal olduğu düşünülürse; 1g karbonhidrat 4 kcal verdiğine göre bir günde yaklaşık 300 g karbonhidrat almamız gerekir. Özellikle beyin dokusu enerji ihtiyacı açısından büyük ölçüde karbonhidratlara bağımlıdır ve kan glukozunun düşüklüğü (hipoglisemi) bu organda ciddi fonksiyon bozukluklarına yol açar (Beynin 1 saattaki glukoz ihtiyacı 6 g dır). Karbohidratlar-Enerji-Yapım Enerji sağlaması dışında karbonhidratlar laktozun, polisakkaridlerin, glikoprotein ve glikolipidlerin yapısına katılırlar. Ayrıca karbonhidrat metabolizmasının zaman zaman protein ve lipid metabolizmaları ile beraber yürüdüğünü, karbonhidratlardan amino asit ve yağ sentezlendiğini, tersine amino asitlerin deaminasyonu ile oluşan karbon iskeletlerinden glukoz sentezlendiğini göreceğiz. Glukoz Glukoz, karbonhidrat metabolizmasının temel maddesidir. Türü ne olursa olsun organizmaya giren her karbonhidrat sonunda glukoza çevrilir. Bu nedenle '' Karbonhidrat metabolizması deyince akla glukoz metabolizması gelir. '' şeklinde düşünmek yerinde bir görüş olur. KARBOHİDRAT METABOLİZMASININ BAŞLICA METABOLİK YOLLARI 1) Glikojenez: Glukozdan glikojen sentezi. 2) Glikojenoliz: Glikojenin yıkılması. Bu olayın karaciğerdeki son ürünü glukoz, kas dokusundaki son ürünü glukoz-6fosfattIr. 3) Glikoliz ( Embden-Meyerhof Yolu) : Glukozun pirüvat veya laktata kadar yıkılması 4) Pirüvat Metabolizması: Pirüvatın asetilKoA ya dönüşümü 5) Trikarboksilik Asit Siklüsü ( Krebs Siklüsü) : Asetil-KoA içindeki asetil kısmının CO2 ye parçalanması ve bu sırada redükte koenzimlerin oluşumu. 6) Pentoz fosfat Yolu: Glukozun bir başka şekilde oksidasyonu ile NADPH ve pentoz sentezi. 7) Glukoneojenezis: Karbonhidrat olmayan maddelerden glukoz sentezi 8) Glukuronik asit Yolu: Glukozdan glukuronik asit sentezi. Karbonhidrat metabolizmasının temel metabolik yolları şekil de topluca gösterilmiştir. KARBOHİDRATLARIN SİNDİRİMİ VE EMİLİMİ Diyette bulunan karbonhidratlar çoğunlukla polisakkarid ve disakkarid (nişasta, laktoz ve sukroz), daha az oranda da monosakkarid (glukoz ve früktoz) formunda Bulunurlar. Karbonhidratların bağırsaktan emilebilmeleri için monosakkarid haline çevrilmeleri şarttır. Sindirim ve Emilim Karbonhidratların parçalanma işlemi ağızdan itibaren başlayabilir. Tükrükte bulunan -amilaz polisakkarid parçalayan bir enzim olmasına rağmen, çiğneme süresinin kısalığı nedeniyle fazla etkin değildir. Midenin asit pH sı bu enzimin daha fazla çalışmasına olanak vermez. Pankreastan çok aktif bir başka amilaz salgılanır. Gerek tükrük gerekse pankreas amilazı, - 1,4 glikozid bağına sahip polisakkaridleri bir bağ atlayarak parçalar ve aktivitesi zincir sonuna yahut dallanma noktasına 2 glukoz ünitesi kalınca durur. - amilaz Böylece - amilaz etkisi sonunda yaklaşık Olarak; % 40 oranında maltoz, % 30 oranında - 1,4 ve - 1,6 bağı içeren - dekstrin % 25 oranında - 1,4 bağı ile bağlanmış 3 glukozlu maltotrioz, % 5 oranında da 4-9 glukoz içeren polisakkarid molekülleri meydana gelir. İnce bağırsak mukozasının fırçamsı kenarı tarafından salgılanan diğer hidrolitik enzimlerle karbohidrat sindirimi sürdürülür ve tamamlanır. Bu enzimlerden - glukozidaz her defasında bir tane olmak üzere 14 bağı parçalar. -dekstrinaz, 16 bağını koparır ve serbest glukoz üniteleri meydana gelir. Yine bağırsak mukozası kaynaklı - galaktozidaz veya laktaz laktozu, sükraz ise sükrozu parçalar ve sonunda glukoz, galaktoz ve früktozdan ibaret monosakkaridler meydana gelir. İncebağırsakta Sindirim Sukraz Sukroz Glukoz + Fruktoz Maltaz Maltoz Glukoz + Glukoz Laktaz Laktoz Glukoz + Galaktoz WWW.biyokimya-pinari.com Monosakkaridlerin ince bağırsaktan emilimleri Bunun için 3 yol vardır. Bunlar aktif transport, taşıyıcı moleküllerle yürütülen kolaylaştırılmış diffüzyon ve basit diffüzyon dur. Monosakkaridlerin transportunda SGLT1 ve GLUT5 isimli glukoz taşıyıcıları (glucose transporter) rol oynar. SGLT1 taşıyıcıları, ince bağırsak hücresinin serozal tarafına yerleşmiş Na+,K+-ATP az sistemine (sodyum pompası) bağımlı olarak çalışır ve monosakkarid, enerji harcanarak düşük konsantrasyondaki bağırsak lumeninden alınır ve yüksek konsantrasyonlu ekstraselüler ortama aktarılır. Glukoz ve galaktoz emilimi tek yönlü olarak çalışan bu yolla gerçekleşir. GLUT5 ise sodyum pompasına bağlı olmaksızın, konsantrasyon farkına dayalı olarak monosakkaridlerin iki yönlü kolaylaştırılmış diffüzyonundan sorumludur. Früktoz ve bir kısım glukoz bu yolla taşınır. Basit diffüzyonda ise taşıyıcı protein yoktur, olayı yönlendiren konsantrasyon gradientidir. Basit diffüzyon glukoz ve früktoz emiliminde kullanılır. GLUKOZ VE HÜCRE Bağırsaklardan emildikten sonra vena porta aracılığı ile sistemik dolaşıma katılan glukoz, galaktoz ve fruktoz hücre düzeyinde tek monosakkarid (glukoz) üzerinden metabolize edilir. Früktozun glukoza dönüşümü bağırsak ve karaciğerde, galaktozun glukoza dönüşümü ise yalnızca karaciğerde gerçekleşir. GLİKOLİZ Glikoz doğada çok yaygın bir monosakkariddir. Hücreler glikozu özel bir yolla yıkar. Canlılar burada açığa çıkan enerjiyi, yaşamsal işlevlerini devam ettirebilmek için kullanırlar. Oksijen kullanmaksızın sitoplazmada gerçekleşen glikozun pirüvik asite(pirüvata) kadar yıkımına “glikoliz” denir. Şeker parçalanması glikoz molekülünün aktivasyonu ile başlar. Glikoz ilk olarak “hekzokinaz” enziminin etkisi altında ATP ile tepkimeye girerek glikoz-6-fosfatı yapar ve kimyasal olarak aktif hale geçer. Glikoz-6-fosfat, glikoza göre enerjice daha zengindir. Glikoz-6-fosfat da fosfo-glikoizomeraz enzimi ile fruktoz-6-fosfata dönüşür. Daha sonra her biri bir enzim tarafından katalizlenen tepkimeler olur. Glikoliz sonucunda 1molekül glikozdan 2 pirüvik asit oluşurken; 4 molekül ADP, ATP’ye çevrilir. Başlangıçta 2 ATP, glikozu, glikoz-6-fosfata ve fruktoz-6-fosfatı, fruktoz-1,6 difosfata çevirmek için kullanıldığından net kazanç 2 ATP’dir. Sonraki tepkimelerde kullanıma hazır 2 molekül de NADH+H+ koenzimi oluşur. C6H12O6+ 2ATP 2C3H4O3(pirüvik asit) + 4H + 2ADP + 2P GLİKOLİZ Glikolizin ilk reaksiyonunda glukoz, glukoz-6-fosfata dönüşmek üzere C-6’da fosfatlanarak sonraki reaksiyonlar için hazırlanır; fosfat donörü ATP’dir: Enzim Heksokinazdır. Glukoz-6-Fosfat, glukoz molekülünü hücre içinde tutan bir tür kapandır ve glukozun hücre dışına çıkışını engeller. Fosforilasyon için 2 ayrı enzimin bulunuşu glukozun karaciğer ve diğer organlar tarafından kullanılış önceliğini belirlemeye yöneliktir. Bu uygulama her iki enzimin Km değerlerinin bilinmesi ile açıklık kazanır. 2) Glukoz-6-fosfatın fruktoz-6-fosfata reversible izomerizasyonunudur. Fosfoheksoz izomeraz (fosfoglukoz izomeraz), katalizler 3) Fruktoz-6-fosfatın fruktoz-1,6bisfosfata fosforilasyonunu, fosfofruktokinaz-1 katalizler; fosfat grubu donörü ATP’dir: 4) Fruktoz-1,6-bisfosfat, fruktoz-1,6-bisfosfat aldolaz (aldolaz) tarafından katalizlenen bir reverzibl aldol kondensasyon reaksiyonunda iki farklı trioz fosfata yıkılır; bir aldoz olan gliseraldehit-3-fosfat ile bir ketoz olan dihidroksiaseton fosfat oluşturur: 5) Dihidroksiaseton fosfat ile gliseraldehit-3-fosfatın karşılıklı dönüşümlerini trioz fosfat izomeraz katalizler: 6) Glikolizin ATP’nin oluştuğu sonuç fazının ilk basamağı, gliseraldehit-3fosfatın 1,3-bisfosfogliserata oksidasyonudur; reaksiyonu, gliseraldehit-3-fosfat dehidrojenaz katalizler: 7) 1,3-bisfosfogliseratın karboksil grubundaki yüksek enerjili fosfat grubu, ATP ve 3fosfogliserat oluşturmak üzere ADP’ye transfer edilir; reaksiyonu fosfogliserat kinaz katalizler: 8) 3-fosfogliserat, 2-fosfogliserata dönüştürülür; reaksiyonu fosfogliserat mutaz katalizler: 9) 2-fosfogliseratın yüksek fosfat grubu transfer potansiyeli olan fosfoenolpirüvata (PEP) dehidrasyonu, enolaz vasıtasıyla katalizlenir: 10) Glikolizin son basamağında fosfoenolpirüvattaki fosfat grubu ADP’ye transfer edilir; ATP ile pirüvat oluşur; reaksiyonu pirüvat kinaz katalizler: 11) Glikolizin düzenlenmesi Louis Pasteur, maya vasıtasıyla glukozun fermantasyonu çalışmaları sırasında, anaerobik şartlar altında glukozun tüketim miktar ve hızının aerobik şartlardakinden çok daha fazla olduğunu buldu. Daha sonraki kas çalışmalarında anaerobik ve aerobik şartlar altındaki glikolizin hızında aynı büyük farklılıklar görüldü. Glikolizin anaerobik şartlarda hızlı aerobik şartlarda yavaş olması Pasteur etkisi olarak bilinir ki bunun biyokimyasal temeli, anaerobik şartlar altında glikolizden üretilen ATP’nin glukozun aerobik şartlar altında tamamen CO2 ve H2O’ya oksidasyonundan üretilen ATP’den çok daha az olmasıdır. Sabit ATP düzeylerini sağlamak için glikolitik yola glukoz akımı düzenlenir. Glikolizin hızında gerekli düzenleme, fosfofruktokinaz-1 (PFK-1) ve pirüvat kinaz enzimlerinin düzenlenmesi vasıtasıyla başarılır. Her iki enzim, ATP üretilmesi ve tüketilmesi arasındaki sellüler dengeyi yansıtan bazı anahtar metabolitlerin konsantrasyonlarındaki değişmelerle allosterik olarak düzenlenirler. Pirüvatın anaerobik ve aerobik şartlarda akıbeti GLİKOLİZİN ENERJİ BİLANÇOSU Glikolizde 2 ATP harcanmaktadır Glikolitik tepkimelerde ise 4 ATP oluşmaktadır. Glikolizin enerji kazancı net 2 ATP olmaktadır. Glikolizin devam edebilmesi için sürekli NAD+ ve ADP sağlanmalıdır. Hücrelerin büyük bir bölümünde O2 kullanılarak oksidatif fosforilasyonla ATP, elde edilmektedir. Cori döngüsü Memeli kaslarında egzersiz sırasında oluşan laktat, kan yoluyla kas hücrelerinden karaciğere taşınır ve hepatik laktat dehidrojenaz etkisiyle pirüvata dönüştürülür. Pirüvat da glukoneojenez yoluyla tekrar glukoza dönüştürülebilir. Glukozun ekstrahepatik dokularda laktata dönüşmesinin ardından laktatın karaciğerde tekrar glukoza dönüşmesi Cori döngüsü olarak bilinir. Glukoz Glukoz NAD+ NADH + H+ 2 Pirüvat NADH + H+ NAD+ GLUKONEOGENEZ 2 ATP GLİKOLİZ NAD+ NADH + H+ 6 ATP 2 Pirüvat NADH + H+ NAD+ 2 Laktat İSKELET KASI 2 Laktat KARACİĞER CORİ DÖNGÜSÜ ALKOL FERMANTASYONU Maya ve diğer mikroorganizmalar glukozu laktattan daha çok etanol ve CO2’e fermente ederler. Alkol fermantasyon olarak adlandırılan bu olay sırasında glukoz, glikoliz yoluyla pirüvata dönüştürülür; pirüvat da iki basamaklı bir proçeste etanol ve CO2’e dönüştürülür. Pirüvatın etanol ve CO2’e dönüştürülmesi proçesinin ilk basamağı pirüvat dekarboksilaz tarafından katalizlenir ve pirüvattan asetaldehit oluşturulur; ikinci basamakta ise asetaldehit, NADH gerektiren alkol dehidrojenaz enzimi tarafından katalizlenen bir reaksiyonda etanole dönüştürülür: Sitrik asit döngüsü, trikarboksilik asit döngüsü (TCA döngüsü), Krebs döngüsü Krebs devri karbonhidratlar, yağlar ve proteinlerin solunumla parçalanması olayında ortak karbon yoludur. Yağ asitleri ve aminoasitler farklı sayıda karbon atomu taşıdıkları için farklı sayıda ATP üretilmesine neden olurlar. Sonuçta oluşan su ve karbondioksit miktarı da farklı olur. Örneğin yağ asitleri az O2, çok H2 taşırlar ve solunum sonucunda az CO2 ve çok H2O oluşur Sitrik asit döngüsü, aerobik metabolizmanın merkezini oluşturur; hücresel solunumda karbonhidrat, yağ ve protein katabolizmasının ortak son ürünü olan asetil-CoA’nın asetil gruplarının oksitlendiği döngüsel olaylar dizisidir: Karbonhidratlardan glikoliz yolunda oluşan pirüvat, aerobik şartlar altında karboksil grubunu CO2 şeklinde kaybetmek suretiyle oksitlenerek asetilCoA’ya dönüşür; pirüvatın oksidatif dekarboksilasyonu olarak bilinen bu reaksiyonu, pirüvat dehidrojenaz enzim kompleksi katalizler: TCA DÖNGÜSÜ Başlıca reaksiyonlar 1) Sitrik asit döngüsünün ilk reaksiyonu, asetil-CoA’nın oksaloasetat ile kondensasyonudur; reaksiyonu sitrat sentaz katalizler ve sitrat oluşur: 2) Akonitaz(akonitat hidrataz) enzimi, sitratın cis-akonitat ara ürünü üzerinden izositrata reversibl dönüşümünü katalizler; bu basamakta dehidrasyon ve hidrasyon reaksiyonları birbirini izler: 3) İzositrat dehidrojenaz enzimi, izositratın oksidatif dekarboksilasyonunu katalizleyerek izositratı α-ketoglutarat ve CO2’e oksitler: 4) α-Ketoglutarat, oksidatif dekarboksilasyona uğrayarak süksinil-CoA ve CO2’e oksitlenir; reaksiyonu αketoglutarat dehidrojenaz enzim kompleksi katalize eder ve NAD+ elektron akseptörü olarak görev görür: 5) Sitrik asit döngüsünde α-ketoglutaratın oksidasyon enerjisi, süksinil-CoA’nın tiyoester bağlarının oluşmasında kullanılmıştır. Sitrik asit döngüsünün sonraki basamağında, süksinil-CoA’nın tiyoester bağlarının yüksek derecede negatif hidroliz standart serbest enerjisi(ΔGoi ≈ −36 kJ/mol), bu bağların yıkılması sırasında salınır ve GTP veya ATP’de bir fosfoanhidrid bağının sentezi için kullanılır. Bu reaksiyon, reversibldir; süksinil-CoA sentetaz veya süksinik tiyokinaz diye adlandırılan enzim tarafından katalizlenir; süksinilCoA’dan süksinat oluşur: 6) Süksinil-CoA’dan oluşan süksinat, flavoprotein süksinat dehidrojenaz vasıtasıyla fumarata okside edilir: 7) Fumaraz (fumarat hidrataz) enzimi vasıtasıyla katalizlenen bir reversibl hidrasyon reaksiyonu sonunda fumarat, L-malata dönüştürülür: 8) Sitrik asit döngüsünün son reaksiyonunda, NAD-bağımlı L-malat dehidrojenaz, L-malatın oksaloasetata oksidasyonunu katalizler: Sitrik asit döngüsü reaksiyonlarından oksaloasetattan sitrat oluşumu ve αketoglutarattan süksinil-KoA oluşumu tek yönlü, diğer reaksiyonlar iki yönlüdür: Sitrik asit döngüsünde elde edilen biyolojik enerji Sitrik asit döngüsünün her dönüşünde üç NADH, bir FADH2 ve bir GTP(veya ATP) ortaya çıkar ve oksidatif dekarboksilasyon reaksiyonlarında iki CO2 serbestleşir: Sitrik asit döngüsünde döngünün her dönüşünde bir ATP molekülü oluşmakla beraber döngüde dört oksidasyon basamağı da solunum zincirine büyük bir elektron akımı sağlar ve böylece sonuç olarak oksidatif fosforilasyon sırasında fazla miktarda ATP oluşmasına yol açar ki solunum zincirinde bir FADH2, 2 ATP oluşmasını sağlar; bir NADH ise 3 ATP oluşmasını sağlar. Bir glukoz molekülünden glikoliz yolunda iki pirüvat oluştuğunu biliyoruz. Bu iki pirüvat da pirüvat dehidrojenaz enzim kompleksi vasıtasıyla asetil-CoA’ya dönüştükten sonra sitrik asit döngüsüne girmektedirler. Sitrik asit döngüsü ve oksidatif fosforilasyon ile bir asetil-CoA molekülünün tam olarak oksitlenmesi sonucunda yaklaşık 12 ATP elde edilmektedir: Bir tek glukoz molekülünün tamamen CO2 ve H2O’ya oksitlenmesi suretiyle net 38 adet ATP kazancı olduğu hesaplanabilir: C6H12O6 + 6O2 ATP (net) Glikoliz Pirüvik asit Krebs Çemberi 6CO2 + H2O + ENERJİ(net 38 ATP) FADH2 NADH2 Net Kazanç 2 ATP __ 2x3 8 ATP __ __ 2x3 6 ATP 2 ATP 2x2 6x3 24 ATP 4 ATP 4 ATP 30 ATP 38 ATP Sitrik asit döngüsünün düzenlenmesi Karbon atomlarının pirüvattan sitrik asit döngüsüne geçişi iki düzeyde sıkı bir şekilde düzenlenir; pirüvat dehidrojenaz kompleksi reaksiyonu vasıtasıyla pirüvatın asetil-CoA’ya dönüşümü basamağı ve sitrat sentaz reaksiyonu vasıtasıyla asetil-CoA’nın döngüye giriş basamağı. Sitrik asit döngüsü, aynı zamanda izositrat dehidrojenaz ve α-ketoglutarat dehidrojenaz reaksiyonlarında düzenlenir: Glioksilat Döngüsü Bitkilerde, bazı omurgasızlarda, E.coli ve maya gibi bazı mikroorganizmalarda asetatın hem yüksek enerji kaynağı olarak hem de karbonhidrat sentezi için fosfoenolpirüvat kaynağı olarak görev görmesi önemlidir. Glioksilat döngüsünde izositrat, izositrat liyaz vasıtasıyla süksinat ve glioksilata yıkılır; daha sonra glioksilat, malat sentaz tarafından katalizlenen bir reaksiyonda malat oluşturmak üzere asetil-CoA ile kondense olur. Glioksilat Döngüsü DİĞER MONOSAKKARİDLERİN METABOLİZMALARI FRUKTOZ Disakkarid Sakkaroz Sakkaraz Fruktoz + Glukoz Karaciğer Besin ve içeceklerde tatlandırıcı ATP Hekzokinaz (Fruktokinaz) Fruktoz 1-fosfat Fruktoz 1,6-bisfosfat Fruktoz 1-fosfat aldolaz Dihidroksiaseton fosfat + Gliseraldehid 3-fosfat Glikoliz 2ATP + 2NADH Fruktoz katabolizmasında fosfofruktokinaz-1 aşaması atlandığı için kontrol mekanizması etkili olmaz. GALAKTOZ Disakkarid Laktoz Laktaz Glukoz+ Galaktoz Galaktokinaz Süt şekeri Galaktoz 1-fosfat UDP-Glukoz Galaktoz 1-fosfat üridil transferaz Glukoz 1-fosfat UDP-Galaktoz UDP-Galaktoz epimeraz UDP-Glukoz Glukoz 1-fosfat Fosfoglukomutaz 2ATP + 2NADH Glikoliz Glukoz 6-fosfat Monosakkarid + Monosakkarid Glikozidik bağ Disakkarid 1,4 -Glikozidik bağ Nişasta Hidroliz D-glukoz D-glukoz Maltoz 1,6 -Glikozidik bağ Nişasta Hidroliz D-glukoz D-glukoz İzomaltoz 1,4 -Glikozidik bağ Selüloz Hidroliz D-glukoz D-glukoz Sellobioz 1,4 -Glikozidik bağ Laktoz Hidroliz D-galaktoz D-glukoz 1,6 -Glikozidik bağ Sakkaroz Hidroliz D-fruktoz D-glukoz POLİSAKKARİDLER Homopolisakkaridler Nişasta Glikojen Glukoz polimeri Glukoz polimeri Bitkisel Hayvansal Düz zincir yapısı Dallı zincir yapısı Heteropolisakkaridler